Phenylethanolamine N-Methyltransferase
2-Hydroxyphenethylamine
Iproniazid
DNA (Cytosine-5-)-Methyltransferase
Dopamine beta-Hydroxylase
Methyltransferases
Tyrosine 3-Monooxygenase
Tetrahydroisoquinolines
Adrenal Medulla
Epinephrine
Adrenal Glands
Aromatic-L-Amino-Acid Decarboxylases
Phenethylamines
Protein Methyltransferases
O(6)-Methylguanine-DNA Methyltransferase
Methylation
Chromaffin Cells
Histone-Lysine N-Methyltransferase
tRNA Methyltransferases
Early Growth Response Protein 1
Protein-Arginine N-Methyltransferases
Gene Expression Regulation, Enzymologic
S-Adenosylmethionine
Protein D-Aspartate-L-Isoaspartate Methyltransferase
DNA Modification Methylases
Site-Specific DNA-Methyltransferase (Adenine-Specific)
Phenylethanolamine N-methyltransferase gene expression. Sp1 and MAZ potential for tissue-specific expression. (1/128)
Phenylethanolamine N-methyltransferase (PNMT) promoter-luciferase reporter gene constructs (pGL3RP863, pGL3RP444, and pGL3RP392) transfected into COS1, RS1, PC12, NIH/3T3, or Neuro2A cells showed the highest basal luciferase activity in the Neuro2A cells. DNase I footprinting with Neuro2A cell nuclear extract identified protected PNMT promoter regions spanning the -168/-165 and -48/-45 base pair Sp1/Egr-1 binding sites. Gel mobility shift assays and transient transfection assays using site-directed mutant PNMT promoter-luciferase reporter gene constructs indicated that the elevated basal luciferase activity in the Neuro2A cells was mediated by Sp-1. Furthermore, activation of the PNMT promoter by Sp1 depends on both its binding affinity for its cognate target sequences and its intracellular concentrations. When Sp1 levels were increased through an expression plasmid, luciferase reporter gene expression rose well beyond basal wild-type levels, even with either Sp1 binding element mutated. Finally, another transcription factor expressed in the Neuro2A cells competes with Sp1 by interacting with DNA sequences 3' to the -48 base pair Sp1 site to prevent Sp1 binding and induction of the PNMT promoter. The DNA consensus sequence, Southwestern analysis, and gel mobility shift assays with antibodies identify MAZ as the competitive factor. These findings suggest that Sp1 may potentially contribute to the tissue-specific expression of the PNMT gene, with the competition between Sp1 and MAZ conferring additional tissue-specific control. (+info)Properties of C1 and other ventrolateral medullary neurones with hypothalamic projections in the rat. (2/128)
1. This study compared (i) the properties of C1 cells with those of neighbouring non-C1 neurones that project to the hypothalamus and (ii) the properties of C1 cells that project to the hypothalamus with those of their medullospinal counterparts. 2. Extracellular recordings were made at three rostrocaudal levels of the ventrolateral medulla (VLM) in alpha-chloralose-anaesthetized, artificially ventilated, paralysed rats. Recorded cells were filled with biotinamide. 3. Level I (0-300 microm behind facial nucleus) contained spontaneously active neurones that were silenced by baro- and cardiopulmonary receptor activation and virtually unaffected by nociceptive stimulation (firing rate altered by < 20 %). These projected either to the cord (type I; 36/39), or to the hypothalamus (type II; 2/39) but rarely to both (1/39). 4. Level II (600-800 microm behind facial nucleus) contained (i) type I neurones (n = 3) (ii) type II neurones (n = 11), (iii) neurones that projected to the hypothalamus and were silenced by baro- and cardiopulmonary receptor activation but activated by strong nociceptive stimulation (type III, n = 2), (iv) non-barosensitive cells activated by weak nociceptive stimulation which projected only to the hypothalamus (type IV, n = 9), (v) cells that projected to the hypothalamus and responded to none of the applied stimuli (type V, n = 7) and (vi) neurones activated by elevating blood pressure which projected neither to the cord nor to the hypothalamus (type VI, n = 4). 5. Level III (1400-1600 microm behind facial motor nucleus) contained all the cell types found at level II except type I. 6. Most of type I and II (17/26) and half of type III cells (4/8) were C1 neurones. Type IV-V were rarely adrenergic (2/12) and type VI were never adrenergic (0/3). 7. All VLM baroinhibited cells project either to the cord or the hypothalamus and virtually all (21/23) C1 cells receive inhibitory inputs from arterial and cardiopulmonary receptors. (+info)Analysis of mice carrying targeted mutations of the glucocorticoid receptor gene argues against an essential role of glucocorticoid signalling for generating adrenal chromaffin cells. (3/128)
Molecular mechanisms underlying the generation of distinct cell phenotypes is a key issue in developmental biology. A major paradigm of determination of neural cell fate concerns the development of sympathetic neurones and neuroendocrine chromaffin cells from a common sympathoadrenal (SA) progenitor cell. Two decades of in vitro experiments have suggested an essential role of glucocorticoid receptor (GR)-mediated signalling in generating chromaffin cells. Targeted mutation of the GR should consequently abolish chromaffin cells. The present analysis of mice lacking GR gene product demonstrates that animals have normal numbers of adrenal chromaffin cells. Moreover, there are no differences in terms of apoptosis and proliferation or in expression of several markers (e.g. GAP43, acetylcholinesterase, adhesion molecule L1) of chromaffin cells in GR-deficient and wild-type mice. However, GR mutant mice lack the adrenaline-synthesizing enzyme PNMT and secretogranin II. Chromaffin cells of GR-deficient mice exhibit the typical ultrastructural features of this cell phenotype, including the large chromaffin granules that distinguish them from sympathetic neurones. Peripherin, an intermediate filament of sympathetic neurones, is undetectable in chromaffin cells of GR mutants. Finally, when stimulated with nerve growth factor in vitro, identical proportions of chromaffin cells from GR-deficient and wild-type mice extend neuritic processes. We conclude that important phenotypic features of chromaffin cells that distinguish them from sympathetic neurones develop normally in the absence of GR-mediated signalling. Most importantly, chromaffin cells in GR-deficient mice do not convert to a neuronal phenotype. These data strongly suggest that the dogma of an essential role of glucocorticoid signalling for the development of chromaffin cells must be abandoned. (+info)Adrenomedullary function is severely impaired in 21-hydroxylase-deficient mice. (4/128)
Deficiency of 21-hydroxylase (21-OH), one of the most common genetic defects in humans, causes low glucocorticoid and mineralocorticoid production by the adrenal cortex, but the effect of this disorder on the adrenomedullary system is unknown. Therefore, we analyzed the development, structure, and function of the adrenal medulla in 21-OH-deficient mice, an animal model resembling human congenital adrenal hyperplasia. Chromaffin cells of 21-OH-deficient mice exhibited ultrastructural features of neuronal transdifferentiation with reduced granules, increased rough endoplasmic reticulum and small neurite outgrowth. Migration of chromaffin cells in the adrenal to form a central medulla was impaired. Expression of phenylethanolamine-N-methyltransferase (PNMT) was reduced to 27 +/- 9% (P<0.05), as determined by quantitative TaqMan polymerase chain reaction, and there was a significant reduction of cells staining positive for PNMT in the adrenal medulla of the 21-OH-deficient mice. Adrenal contents of epinephrine were decreased to 30 +/- 2% (P<0. 01) whereas norepinephrine and dopamine levels were reduced to 57 +/- 4% (P<0.01) and 50 +/- 9% (P<0.05), respectively. 21-OH-deficient mice demonstrate severe adrenomedullary dysfunction, with alterations in chromaffin cell migration, development, structure, and catecholamine synthesis. This hitherto unrecognized mechanism may contribute to the frequent clinical, mental, and therapeutic problems encountered in humans with this genetic disease. (+info)Acetylcholinesterase activity, and neurofilament protein, and catecholamine synthesizing enzymes immunoreactivities in the mouse adrenal gland during postnatal development. (5/128)
The present study showed the acetylcholinesterase (AChE) activity, and neurofilament protein (NFP), catecholamine-synthesizing enzymes, dopamine beta-hydroxylase (DBH) and phenylethanolamine N-methyltransferase (PNMT) immunoreactivities in the mouse adrenal gland during postnatal development. From birth to postnatal-1-day, AChE activity was weakly and diffusely found in some medullary cells and in very few nerve fibers whereas strong NFP immunoreactivity was seen in a few ganglion cells and in remarkably numerous nerve fibers in the medulla. Almost all meduallary cells were reactive for both DBH and PNMT during this period. From postnatal-2- or -3-day to postnatal-1-week, strong AChE activity was observed in a few large ganglion cells, but the reaction was weak in clusters of chromaffin cells, and the number of strong AChE-active nerve fibers in the medulla was rapidly increased. From postnatal-2-day onwards, the number of NFP-immunoreactive nerve fibers in the medulla were remarkably numerous. Numerous chromaffin cells were reactive for both DBH and PNMT whereas some chromaffin cells were reactive for only DBH from postnatal-2-day onwards. These results suggest that drastic changes such as an increase of acetylcholine in the nerve fibers, differentiation of noradrenaline and adrenaline cells of the medulla may occur during this period. From postnatal-2-week to postnatal-3-week, weak AChE activity was seen in the clusters of several chromaffin cells and a few ganglion cells, and the number of AChE-active nerve fibers in the medulla was gradually increased. From postnatal-4-week to postnatal-8-week (adult), the distribution and frequency of AChE activity in the adrenal gland were similar to those at postnatal-3-week. In the adult, AChE activity was weakly seen in the clusters of several chromaffin cells showing noradrenaline fluorescence in the adrenal medulla. The noradrenaline cells were contacted by denser AChE-reactive nerve fibers than adrenaline cells. These results suggest that the development of cholinergic nervous system in the mouse adrenal medulla may be completed by postnatal-3-week. (+info)Lesions of the C1 catecholaminergic neurons of the ventrolateral medulla in rats using anti-DbetaH-saporin. (6/128)
Phenylethanolamine-N-methyltransferase (PNMT)-containing neurons in the rostral ventrolateral medulla (RVLM) are believed to play a role in cardiovascular regulation. To determine whether injection of anti-dopamine beta-hydroxylase (DbetaH)-saporin directly into the RVLM in rats could selectively destroy these cells and thereby provide an approach for evaluating their role in cardiovascular regulation, we studied rats 2 wk after unilateral injection of 21 ng anti-DbetaH-saporin into the RVLM. There was an approximately 90% reduction in the number of PNMT-positive neurons in the RVLM, although the number of non-C1, spinally projecting barosensitive neurons of this area was not altered. The A5 cell group was the only other population of DbetaH-containing cells that was significantly depleted. The depressor response evoked by injection of tyramine into the RVLM was abolished by prior injection of toxin. The pressor response evoked by injection of glutamate into the RVLM was attenuated ipsilateral to the toxin injection but was potentiated contralateral to the toxin injection. Thus anti-DbetaH-saporin can be used to make selective lesions of PNMT-containing cells, allowing for the evaluation of their role in cardiovascular regulation. (+info)Nicotine infusion modulates immobilization stress-triggered induction of gene expression of rat catecholamine biosynthetic enzymes. (7/128)
The relationship between nicotine and stress is complex and paradoxical. Although people claim they smoke because it relaxes them, nicotine can trigger some of the effects observed with stress, including the release and synthesis of the catecholamines and their biosynthetic enzymes. This study examined one aspect of this confusing relationship between nicotine and stress. Multiple injections of nicotine bitartrate (5 mg/kg) elevated mRNA levels for the catecholamine biosynthetic enzymes, tyrosine hydroxylase (TH), dopamine beta-hydroxylase (DBH), and phenylethanolamine N-methyltransferase, and of preproneuropeptide Y in rat adrenal medulla more than did 1 mg/kg of nicotine bitartrate. In the locus ceruleus, substantia nigra, and ventral tegmental area both doses equally induced TH mRNA levels. Nicotine infusion (15 mg/kg/day) did not affect adrenal mRNA levels for any of the genes of interest and did not increase plasma corticosterone levels. However, in rats pre-exposed to nicotinic infusions, the response to a single immobilization (IMO) stress was markedly attenuated with respect to changes in adrenomedullary TH, DBH, and phenylethanolamine N-methyltransferase mRNA levels and in c-Fos protein levels. In the central nervous system, the chronic infusion of nicotine prevented the induction of TH mRNA by repeated IMO stress in the ventral tegmental area (but not in substantia nigra) and of DBH mRNA by single IMO in the locus ceruleus. These findings may explain some of the complex interactions between stress and exposure to nicotine. (+info)Actions of hypoxia on catecholamine synthetic enzyme mRNA expression before and after development of adrenal innervation in the sheep fetus. (8/128)
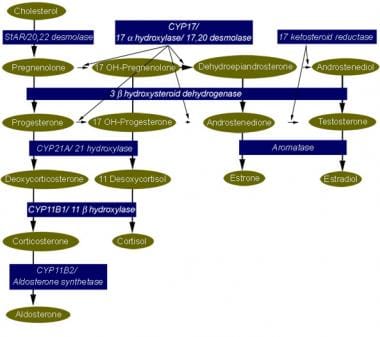
We have investigated adrenal mRNA expression of the catecholamine synthetic enzymes tyrosine hydroxylase (TH) and phenylethanolamine N-methyltransferase (PNMT) following acute hypoxia in fetal sheep before (< 105 days gestation, n = 20) and after (> 125 days gestation, n = 20) the development of adrenal innervation and following pretreatment with the nicotinic receptor anatgonist hexamethonium (n = 12). Total RNA was extracted from fetal adrenal glands collected at specific time points at 3-20 h after the onset of either hypoxia ( approximately 50% reduction in fetal arterial oxygen saturation (SO2) for 30 min), or normoxia. Before 105 days, there was a decrease in adrenal TH mRNA expression at 20 h after hypoxia and adrenal TH mRNA expression was directly related to the changes in arterial PO2 measured during normoxia and hypoxia. After 125 days, adrenal TH mRNA levels were suppressed for up to 12 h following hypoxia. In both age groups, adrenal PNMT mRNA expression increased at 3-5 h after hypoxia and was inversely related to the changes in fetal arterial PO2 during normoxia or hypoxia. After 125 days, the administration of hexamethonium (25 mg kg(-1), I.V.) reduced TH mRNA but not PNMT mRNA expression after normoxia. After hexamethonium pretreatment, there was no significant change in either adrenal TH or PNMT mRNA expression following hypoxia. We conclude that acute hypoxia differentially regulates adrenal TH and PNMT mRNA expression in the fetal sheep both before and after the development of adrenal innervation. After the development of adrenal innervation, however, the effect of acute hypoxia upon adrenal TH and PNMT mRNA expression is dependent upon neurogenic input acting via nicotinic receptors. (+info)Phenylethanolamine N-Methyltransferase (PNMT) is a enzyme that plays a crucial role in the synthesis of epinephrine (also known as adrenaline). It catalyzes the transfer of a methyl group from S-adenosylmethionine to the nitrogen atom of the amine group of normetanephrine, resulting in the formation of epinephrine.
PNMT is primarily found in the chromaffin cells of the adrenal medulla, where it is responsible for the final step in the biosynthesis of epinephrine. The activity of PNMT is regulated by several factors, including glucocorticoids, which increase its expression and activity, leading to an elevation in epinephrine levels.
Epinephrine is a hormone and neurotransmitter that plays a critical role in the body's response to stress, preparing it for the "fight or flight" response by increasing heart rate, blood pressure, and respiration, among other effects.
2-Hydroxyphenethylamine is a chemical compound that is classified as a phenylethylamine and a hydroxyamphetamine. It is a secondary amine with a hydroxy group attached to the benzene ring, specifically at the 2-position. This compound is a derivative of phenethylamine by the replacement of one hydrogen atom by a hydroxy group.
It is worth noting that 2-hydroxyphenethylamine itself does not have a recognized medical definition or specific clinical relevance. However, it may be encountered in the context of biochemistry, pharmacology, or forensic science. Like other phenylethylamines, it has structural similarity to certain neurotransmitters and drugs, and therefore may have potential pharmacological activity.
Iproniazid is a monoamine oxidase inhibitor (MAOI) drug that was initially used as an antitubercular agent but later found to have antidepressant properties. It works by blocking the breakdown of certain neurotransmitters, such as serotonin and dopamine, in the brain which helps to elevate mood and improve symptoms of depression. However, its use is limited due to the risk of serious side effects, including hypertensive crisis and serotonin syndrome, when taken with certain foods or other medications.
Dopamine beta-hydroxylase (DBH) is an enzyme that plays a crucial role in the synthesis of catecholamines, which are important neurotransmitters and hormones in the human body. Specifically, DBH converts dopamine into norepinephrine, another essential catecholamine.
DBH is primarily located in the adrenal glands and nerve endings of the sympathetic nervous system. It requires molecular oxygen, copper ions, and vitamin C (ascorbic acid) as cofactors to perform its enzymatic function. Deficiency or dysfunction of DBH can lead to various medical conditions, such as orthostatic hypotension and neuropsychiatric disorders.
Methyltransferases are a class of enzymes that catalyze the transfer of a methyl group (-CH3) from a donor molecule to an acceptor molecule, which is often a protein, DNA, or RNA. This transfer of a methyl group can modify the chemical and physical properties of the acceptor molecule, playing a crucial role in various cellular processes such as gene expression, signal transduction, and DNA repair.
In biochemistry, methyltransferases are classified based on the type of donor molecule they use for the transfer of the methyl group. The most common methyl donor is S-adenosylmethionine (SAM), a universal methyl group donor found in many organisms. Methyltransferases that utilize SAM as a cofactor are called SAM-dependent methyltransferases.
Abnormal regulation or function of methyltransferases has been implicated in several diseases, including cancer and neurological disorders. Therefore, understanding the structure, function, and regulation of these enzymes is essential for developing targeted therapies to treat these conditions.
Tyrosine 3-Monooxygenase (also known as Tyrosinase or Tyrosine hydroxylase) is an enzyme that plays a crucial role in the synthesis of catecholamines, which are neurotransmitters and hormones in the body. This enzyme catalyzes the conversion of the amino acid L-tyrosine to 3,4-dihydroxyphenylalanine (L-DOPA) by adding a hydroxyl group to the 3rd carbon atom of the tyrosine molecule.
The reaction is as follows:
L-Tyrosine + O2 + pterin (co-factor) -> L-DOPA + pterin (oxidized) + H2O
This enzyme requires molecular oxygen and a co-factor such as tetrahydrobiopterin to carry out the reaction. Tyrosine 3-Monooxygenase is found in various tissues, including the brain and adrenal glands, where it helps regulate the production of catecholamines like dopamine, norepinephrine, and epinephrine. Dysregulation of this enzyme has been implicated in several neurological disorders, such as Parkinson's disease.
Nordefrin is not typically used as a medical diagnosis or treatment, but it is a medication that contains the active ingredient Noradrenaline (also known as Norepinephrine) which is a naturally occurring hormone and neurotransmitter in the human body.
Noradrenaline is a potent vasoconstrictor, increasing blood pressure and improving blood flow to vital organs such as the heart and brain. It also acts as a bronchodilator, opening up the airways in the lungs. Nordefrin is used as a medication to treat hypotension (low blood pressure) and shock, particularly in cases where other treatments have been ineffective.
It's important to note that Nordefrin should only be administered under the supervision of a healthcare professional, as it can have serious side effects if not used correctly.
Tetrahydroisoquinolines (TIQs) are not a medical condition, but rather a class of organic compounds that have been studied in the field of medicine and neuroscience. TIQs are naturally occurring substances found in various foods, beverages, and plants, as well as produced endogenously in the human body. They have been shown to have various pharmacological activities, including acting as weak psychoactive agents, antioxidants, and inhibitors of certain enzymes. Some TIQs have also been implicated in the pathophysiology of certain neurological disorders such as Parkinson's disease. However, more research is needed to fully understand their roles and potential therapeutic applications.
The adrenal medulla is the inner part of the adrenal gland, which is located on top of the kidneys. It is responsible for producing and releasing hormones such as epinephrine (also known as adrenaline) and norepinephrine (also known as noradrenaline). These hormones play a crucial role in the body's "fight or flight" response, preparing the body for immediate action in response to stress.
Epinephrine increases heart rate, blood pressure, and respiratory rate, while also increasing blood flow to muscles and decreasing blood flow to the skin and digestive system. Norepinephrine has similar effects but is generally less potent than epinephrine. Together, these hormones help to prepare the body for physical activity and increase alertness and focus.
Disorders of the adrenal medulla can lead to a variety of symptoms, including high blood pressure, rapid heart rate, anxiety, and tremors. Some conditions that affect the adrenal medulla include pheochromocytoma, a tumor that causes excessive production of epinephrine and norepinephrine, and neuroblastoma, a cancerous tumor that arises from immature nerve cells in the adrenal gland.
Epinephrine, also known as adrenaline, is a hormone and a neurotransmitter that is produced in the body. It is released by the adrenal glands in response to stress or excitement, and it prepares the body for the "fight or flight" response. Epinephrine works by binding to specific receptors in the body, which causes a variety of physiological effects, including increased heart rate and blood pressure, improved muscle strength and alertness, and narrowing of the blood vessels in the skin and intestines. It is also used as a medication to treat various medical conditions, such as anaphylaxis (a severe allergic reaction), cardiac arrest, and low blood pressure.
Catecholamines are a group of hormones and neurotransmitters that are derived from the amino acid tyrosine. The most well-known catecholamines are dopamine, norepinephrine (also known as noradrenaline), and epinephrine (also known as adrenaline). These hormones are produced by the adrenal glands and are released into the bloodstream in response to stress. They play important roles in the "fight or flight" response, increasing heart rate, blood pressure, and alertness. In addition to their role as hormones, catecholamines also function as neurotransmitters, transmitting signals in the nervous system. Disorders of catecholamine regulation can lead to a variety of medical conditions, including hypertension, mood disorders, and neurological disorders.
The adrenal glands are a pair of endocrine glands that are located on top of the kidneys. Each gland has two parts: the outer cortex and the inner medulla. The adrenal cortex produces hormones such as cortisol, aldosterone, and androgens, which regulate metabolism, blood pressure, and other vital functions. The adrenal medulla produces catecholamines, including epinephrine (adrenaline) and norepinephrine (noradrenaline), which help the body respond to stress by increasing heart rate, blood pressure, and alertness.
Aromatic-L-amino-acid decarboxylases (ALADs) are a group of enzymes that play a crucial role in the synthesis of neurotransmitters and biogenic amines in the body. These enzymes catalyze the decarboxylation of aromatic L-amino acids, such as L-dopa, L-tryptophan, and L-phenylalanine, to produce corresponding neurotransmitters or biogenic amines, including dopamine, serotonin, and histamine, respectively.
There are two main types of ALADs in humans: dopa decarboxylase (DDC) and tryptophan hydroxylase (TPH). DDC is responsible for the conversion of L-dopa to dopamine, which is a crucial neurotransmitter involved in movement regulation. TPH, on the other hand, catalyzes the rate-limiting step in serotonin synthesis by converting L-tryptophan to 5-hydroxytryptophan (5-HTP), which is then converted to serotonin by another enzyme called aromatic amino acid decarboxylase.
Deficiencies or mutations in ALADs can lead to various neurological and psychiatric disorders, such as Parkinson's disease, dopa-responsive dystonia, and depression. Therefore, understanding the function and regulation of ALADs is essential for developing effective therapies for these conditions.
Phenethylamines are a class of organic compounds that share a common structural feature, which is a phenethyl group (a phenyl ring bonded to an ethylamine chain). In the context of pharmacology and neuroscience, "phenethylamines" often refers to a specific group of psychoactive drugs, including stimulants like amphetamine and mescaline, a classic psychedelic. These compounds exert their effects by modulating the activity of neurotransmitters in the brain, such as dopamine, norepinephrine, and serotonin. It is important to note that many phenethylamines have potential for abuse and are controlled substances.
Protein methyltransferases (PMTs) are a family of enzymes that transfer methyl groups from a donor, such as S-adenosylmethionine (SAM), to specific residues on protein substrates. This post-translational modification plays a crucial role in various cellular processes, including epigenetic regulation, signal transduction, and protein stability.
PMTs can methylate different amino acid residues, such as lysine, arginine, and histidine, on proteins. The methylation of these residues can lead to changes in the charge, hydrophobicity, or interaction properties of the target protein, thereby modulating its function.
For example, lysine methyltransferases (KMTs) are a subclass of PMTs that specifically methylate lysine residues on histone proteins, which are the core components of nucleosomes in chromatin. Histone methylation can either activate or repress gene transcription, depending on the specific residue and degree of methylation.
Protein arginine methyltransferases (PRMTs) are another subclass of PMTs that methylate arginine residues on various protein substrates, including histones, transcription factors, and RNA-binding proteins. Arginine methylation can also affect protein function by altering its interaction with other molecules or modulating its stability.
Overall, protein methyltransferases are essential regulators of cellular processes and have been implicated in various diseases, including cancer, neurodegenerative disorders, and cardiovascular diseases. Therefore, understanding the mechanisms and functions of PMTs is crucial for developing novel therapeutic strategies to target these diseases.
Methylation, in the context of genetics and epigenetics, refers to the addition of a methyl group (CH3) to a molecule, usually to the nitrogenous base of DNA or to the side chain of amino acids in proteins. In DNA methylation, this process typically occurs at the 5-carbon position of cytosine residues that precede guanine residues (CpG sites) and is catalyzed by enzymes called DNA methyltransferases (DNMTs).
DNA methylation plays a crucial role in regulating gene expression, genomic imprinting, X-chromosome inactivation, and suppression of repetitive elements. Hypermethylation or hypomethylation of specific genes can lead to altered gene expression patterns, which have been associated with various human diseases, including cancer.
In summary, methylation is a fundamental epigenetic modification that influences genomic stability, gene regulation, and cellular function by introducing methyl groups to DNA or proteins.
Chromaffin cells are specialized neuroendocrine cells that are responsible for the synthesis and release of catecholamines, which are hormones such as adrenaline (epinephrine) and noradrenaline (norepinephrine). These cells are located in the medulla of the adrenal gland and in some autonomic ganglia outside the central nervous system. Chromaffin cells contain secretory granules that stain brown with chromium salts, hence their name. They play a crucial role in the body's response to stress by releasing catecholamines into the bloodstream, which helps prepare the body for the "fight or flight" response.
Histone-Lysine N-Methyltransferase is a type of enzyme that transfers methyl groups to specific lysine residues on histone proteins. These histone proteins are the main protein components of chromatin, which is the complex of DNA and proteins that make up chromosomes.
Histone-Lysine N-Methyltransferases play a crucial role in the regulation of gene expression by modifying the structure of chromatin. The addition of methyl groups to histones can result in either the activation or repression of gene transcription, depending on the specific location and number of methyl groups added.
These enzymes are important targets for drug development, as their dysregulation has been implicated in various diseases, including cancer. Inhibitors of Histone-Lysine N-Methyltransferases have shown promise in preclinical studies for the treatment of certain types of cancer.
tRNA (transfer RNA) methyltransferases are a group of enzymes that catalyze the transfer of a methyl group (-CH3) to specific positions on the tRNA molecule. These enzymes play a crucial role in modifying and regulating tRNA function, stability, and interaction with other components of the translation machinery during protein synthesis.
The addition of methyl groups to tRNAs can occur at various sites, including the base moieties of nucleotides within the anticodon loop, the TψC loop, and the variable region. These modifications help maintain the structural integrity of tRNA molecules, enhance their ability to recognize specific codons during translation, and protect them from degradation by cellular nucleases.
tRNA methyltransferases are classified based on the type of methylation they catalyze:
1. N1-methyladenosine (m1A) methyltransferases: These enzymes add a methyl group to the N1 position of adenosine residues in tRNAs. An example is TRMT6/TRMT61A, which methylates adenosines at position 58 in human tRNAs.
2. N3-methylcytosine (m3C) methyltransferases: These enzymes add a methyl group to the N3 position of cytosine residues in tRNAs. An example is Dnmt2, which methylates cytosines at position 38 in various organisms.
3. N7-methylguanosine (m7G) methyltransferases: These enzymes add a methyl group to the N7 position of guanosine residues in tRNAs, primarily at position 46 within the TψC loop. An example is Trm8/Trm82, which catalyzes this modification in yeast and humans.
4. 2'-O-methylated nucleotides (Nm) methyltransferases: These enzymes add a methyl group to the 2'-hydroxyl group of ribose sugars in tRNAs, which can occur at various positions throughout the molecule. An example is FTSJ1, which methylates uridines at position 8 in human tRNAs.
5. Pseudouridine (Ψ) synthases: Although not technically methyltransferases, pseudouridine synthases catalyze the isomerization of uridine to pseudouridine, which can enhance tRNA stability and function. An example is Dyskerin (DKC1), which introduces Ψ at various positions in human tRNAs.
These enzymes play crucial roles in modifying tRNAs, ensuring proper folding, stability, and function during translation. Defects in these enzymes can lead to various diseases, including neurological disorders, cancer, and premature aging.
Early Growth Response Protein 1 (EGR1) is a transcription factor that belongs to the EGR family of proteins, which are also known as zinc finger transcription factors. EGR1 plays crucial roles in various biological processes, including cell proliferation, differentiation, and apoptosis. It regulates gene expression by binding to specific DNA sequences in the promoter regions of target genes.
EGR1 is rapidly induced in response to a variety of stimuli, such as growth factors, neurotransmitters, and stress signals. Once induced, EGR1 modulates the transcription of downstream target genes involved in different signaling pathways, such as mitogen-activated protein kinase (MAPK), phosphatidylinositol 3-kinase (PI3K), and nuclear factor kappa B (NF-κB) pathways.
EGR1 has been implicated in several physiological and pathological processes, including development, learning and memory, neurodegeneration, and cancer. In the context of cancer, EGR1 can act as a tumor suppressor or an oncogene, depending on the cellular context and the specific target genes it regulates.
Protein-Arginine N-Methyltransferases (PRMTs) are a group of enzymes that catalyze the transfer of methyl groups from S-adenosylmethionine to specific arginine residues in proteins, leading to the formation of N-methylarginines. This post-translational modification plays a crucial role in various cellular processes such as signal transduction, DNA repair, and RNA processing. There are nine known PRMTs in humans, which can be classified into three types based on the type of methylarginine produced: Type I (PRMT1, 2, 3, 4, 6, and 8) produce asymmetric dimethylarginines, Type II (PRMT5 and 9) produce symmetric dimethylarginines, and Type III (PRMT7) produces monomethylarginine. Aberrant PRMT activity has been implicated in several diseases, including cancer and neurological disorders.
Gene expression regulation, enzymologic refers to the biochemical processes and mechanisms that control the transcription and translation of specific genes into functional proteins or enzymes. This regulation is achieved through various enzymatic activities that can either activate or repress gene expression at different levels, such as chromatin remodeling, transcription factor activation, mRNA processing, and protein degradation.
Enzymologic regulation of gene expression involves the action of specific enzymes that catalyze chemical reactions involved in these processes. For example, histone-modifying enzymes can alter the structure of chromatin to make genes more or less accessible for transcription, while RNA polymerase and its associated factors are responsible for transcribing DNA into mRNA. Additionally, various enzymes are involved in post-transcriptional modifications of mRNA, such as splicing, capping, and tailing, which can affect the stability and translation of the transcript.
Overall, the enzymologic regulation of gene expression is a complex and dynamic process that allows cells to respond to changes in their environment and maintain proper physiological function.
S-Adenosylmethionine (SAMe) is a physiological compound involved in methylation reactions, transulfuration pathways, and aminopropylation processes in the body. It is formed from the coupling of methionine, an essential sulfur-containing amino acid, and adenosine triphosphate (ATP) through the action of methionine adenosyltransferase enzymes.
SAMe serves as a major methyl donor in various biochemical reactions, contributing to the synthesis of numerous compounds such as neurotransmitters, proteins, phospholipids, nucleic acids, and other methylated metabolites. Additionally, SAMe plays a crucial role in the detoxification process within the liver by participating in glutathione production, which is an important antioxidant and detoxifying agent.
In clinical settings, SAMe supplementation has been explored as a potential therapeutic intervention for various conditions, including depression, osteoarthritis, liver diseases, and fibromyalgia, among others. However, its efficacy remains a subject of ongoing research and debate within the medical community.
DNA cytosine methylases are a type of enzyme that catalyze the transfer of a methyl group (-CH3) to the carbon-5 position of the cytosine ring in DNA, forming 5-methylcytosine. This process is known as DNA methylation and plays an important role in regulating gene expression, genomic imprinting, X-chromosome inactivation, and suppression of transposable elements in eukaryotic organisms.
In mammals, the most well-studied DNA cytosine methylases are members of the DNMT (DNA methyltransferase) family, including DNMT1, DNMT3A, and DNMT3B. DNMT1 is primarily responsible for maintaining existing methylation patterns during DNA replication, while DNMT3A and DNMT3B are involved in establishing new methylation patterns during development and differentiation.
Abnormal DNA methylation patterns have been implicated in various diseases, including cancer, where global hypomethylation and promoter-specific hypermethylation can contribute to genomic instability, chromosomal aberrations, and silencing of tumor suppressor genes.
Protein D-aspartate-L-isoaspartate methyltransferase (PCMT or PRMT5) is an enzyme that catalyzes the transfer of a methyl group from S-adenosylmethionine to the side chain nitrogen atom of a specific aspartate or glutamate residue on protein substrates. This enzyme plays a crucial role in the maintenance of protein structure and function by correcting the spontaneous deamidation of asparagine and isomerization of aspartate to isoaspartate residues, which can lead to protein aggregation and loss of function. PCMT also regulates various cellular processes, including transcription, RNA processing, DNA damage repair, and signal transduction, by modifying the activity or localization of its target proteins.
DNA modification methylases are a type of enzyme that catalyze the transfer of methyl groups (-CH3) to specific nucleotides in DNA, usually cytosine or adenine residues. This process is known as DNA methylation and is an important epigenetic mechanism that regulates gene expression, genome stability, and other cellular processes.
There are several types of DNA modification methylases, including:
1. Cytosine-5 methyltransferases (CNMTs or DNMTs): These enzymes catalyze the transfer of a methyl group to the fifth carbon atom of cytosine residues in DNA, forming 5-methylcytosine (5mC). This is the most common type of DNA methylation and plays a crucial role in gene silencing, X-chromosome inactivation, and genomic imprinting.
2. N6-adenine methyltransferases (MTases): These enzymes catalyze the transfer of a methyl group to the sixth nitrogen atom of adenine residues in DNA, forming N6-methyladenine (6mA). This type of DNA methylation is less common than 5mC but has been found to be involved in various cellular processes, such as transcriptional regulation and DNA repair.
3. GpC methyltransferases: These enzymes catalyze the transfer of a methyl group to the second carbon atom of guanine residues in DNA, forming N4-methylcytosine (4mC). This type of DNA methylation is relatively rare and has been found mainly in prokaryotic genomes.
Dysregulation of DNA modification methylases has been implicated in various diseases, including cancer, neurological disorders, and immunological diseases. Therefore, understanding the function and regulation of these enzymes is essential for developing novel therapeutic strategies to treat these conditions.
DNA methylation is a process by which methyl groups (-CH3) are added to the cytosine ring of DNA molecules, often at the 5' position of cytospine phosphate-deoxyguanosine (CpG) dinucleotides. This modification is catalyzed by DNA methyltransferase enzymes and results in the formation of 5-methylcytosine.
DNA methylation plays a crucial role in the regulation of gene expression, genomic imprinting, X chromosome inactivation, and suppression of transposable elements. Abnormal DNA methylation patterns have been associated with various diseases, including cancer, where tumor suppressor genes are often silenced by promoter methylation.
In summary, DNA methylation is a fundamental epigenetic modification that influences gene expression and genome stability, and its dysregulation has important implications for human health and disease.
 Phenylethanolamine N-methyltransferase
Phenylethanolamine N-methyltransferase
Methyltransferase
Phenylethanolamine
RU-28362
Halostachine
Adrenaline
History of catecholamine research
3,4-Dichloroamphetamine
Jennifer L. Martin
Adrenal gland
Dopamine
Dextroamphetamine
Amphetamine
Catecholamine
Reward dependence
Adderall
Norepinephrine
Adrenergic cell group C3
Adrenergic cell group C1
N-Methyltyramine
Sympathoadrenal system
Phenethylamine
N-methyltransferase
N-Methylphenethylamine
Biogenic amine
Synthetic drug
List of EC numbers (EC 2)
Trace amine
Synephrine
Monoamine neurotransmitter
Phenylethanolamine N-methyltransferase - Wikipedia
Genistein - Wikipedia
 PDB 1N7J | Chain THE STRUCTURE OF PHENYLETHANOLAMINE N-METHYLTRANSFERASE IN COMPLEX WITH S-ADENOSYLHOMOCYSTEINE AND AN...
PDB 1N7J | Chain THE STRUCTURE OF PHENYLETHANOLAMINE N-METHYLTRANSFERASE IN COMPLEX WITH S-ADENOSYLHOMOCYSTEINE AND AN...
 metabolic pathway pertinent to the brain - Ontology Report - Rat Genome Database
metabolic pathway pertinent to the brain - Ontology Report - Rat Genome Database
 SCOPe 2.08: Domain d2g71a2: 2g71 A:21-280
SCOPe 2.08: Domain d2g71a2: 2g71 A:21-280
SCOPe 2.07: Domain d2g71b2: 2g71 B:14-280
BindingDB BDBM28422 (2S)-2-amino-4-({[(2S,3S,4R,5R)-5-(6-amino-9H-purin-9-yl)-3,4-dihydroxyoxolan-2-yl]methyl}(methyl...
 1,2,3,4-Tetrahydroisoquinoline 95 91-21-4
1,2,3,4-Tetrahydroisoquinoline 95 91-21-4
 Muscarine | Harvard Catalyst Profiles | Harvard Catalyst
Muscarine | Harvard Catalyst Profiles | Harvard Catalyst
 Neurotransmitters - Proteopedia, life in 3D
Neurotransmitters - Proteopedia, life in 3D
 Phenylketonuria - New World Encyclopedia
Phenylketonuria - New World Encyclopedia
 Plus it
Plus it
 Native MS Bibliography | Bruker
Native MS Bibliography | Bruker
 Figures and data in Succinate mediates inflammation-induced adrenocortical dysfunction | eLife
Figures and data in Succinate mediates inflammation-induced adrenocortical dysfunction | eLife
 Professor Brett Collins - Queensland Brain Institute - University of Queensland
Professor Brett Collins - Queensland Brain Institute - University of Queensland
 QuartataWeb 1.0
QuartataWeb 1.0
All Targets-AlzheimerNet
 Potential Biomarkers of Metastasizing Paragangliomas and Pheochromocytomas | Encyclopedia MDPI
Potential Biomarkers of Metastasizing Paragangliomas and Pheochromocytomas | Encyclopedia MDPI
Ronald de Krijger - Onderzoeksoutput - Prinses Máxima Centrum
Microsoft word - dear author.doc
 ADA2A RAT - Alpha-2a Adrenergic Receptor, Rat | ZINC Is Not Commercial - A database of commercially-available compounds
ADA2A RAT - Alpha-2a Adrenergic Receptor, Rat | ZINC Is Not Commercial - A database of commercially-available compounds
 NIKOLSKY BREAST CANCER 17Q11 Q21 AMPLICON
NIKOLSKY BREAST CANCER 17Q11 Q21 AMPLICON
June | 2020 | Thrombin Inhibitors
 Tyrosine - Nootropics Expert
Tyrosine - Nootropics Expert
 human biology - What is the relationship between Homocysteine and Norepinephrine metabolic cycles? - Biology Stack Exchange
human biology - What is the relationship between Homocysteine and Norepinephrine metabolic cycles? - Biology Stack Exchange
STU Health & Medical Enzyme Conversion Description Discussion Nursing Assignment Help
 The Best Damn Cortisol Article Ever - Bigger Stronger Leaner - COMMUNITY - T NATION
The Best Damn Cortisol Article Ever - Bigger Stronger Leaner - COMMUNITY - T NATION
 Pages that link to "Phenelzine" - wikidoc
Pages that link to "Phenelzine" - wikidoc
 Pesquisa | Portal Regional da BVS
Pesquisa | Portal Regional da BVSPNMT12
- Phenylethanolamine N-methyltransferase (PNMT) is an enzyme found primarily in the adrenal medulla that converts norepinephrine (noradrenaline) to epinephrine (adrenaline). (wikipedia.org)
- Several features of the structure like this folding lip suggest that PNMT is a recent adaptation to the catecholamine synthesizing enzyme family, evolving later than COMT, but before other methyltransferases like GNMT. (wikipedia.org)
- While PNMT methylates norepinephrine into the active compound epinephrine, norepinephrine can also be methylated by catechol-O-methyl transferase (COMT), another methyltransferase which adds a methyl group in a different location, in turn producing the inactive compound metanephrine. (wikipedia.org)
- Epinephrine then is synthesized from norepinephrine via methylation of the primary distal amine of norepinephrine by phenylethanolamine N-methyltransferase (PNMT). (newworldencyclopedia.org)
- The activity of phenylethanolamine-N-methyl transferase (PNMT) in the adrenal medulla declines markedly after hypophysectomy. (aspetjournals.org)
- mRNA expression of tyrosine hydroxylase ( Th ) ( D ) and phenylethanolamine N -methyltransferase ( Pnmt ) ( E ) in medulla, cortex, and CD31 - CD45 - cells (ACC) (n=4 mice per group). (elifesciences.org)
- Norepinephrine is converted to epinephrine by phenylethanolamine N -methyltransferase (PNMT). (stackexchange.com)
- In mammals PNMT is just one member of a large family of methyltransferases which use S -adenosylmethionine (AdoMet) as a cofactor to provide a methyl group. (stackexchange.com)
- The conversion of norepinephrine to epinephrine requires the enzyme phenylethanolamine-N-methyltransferase (PNMT). (speedyhomeworktutors.com)
- Cortisol increases the level and activity of an enzyme called Phenylethanolamine-N-methyltransferase (PNMT). (t-nation.com)
- Phase V of the PFS Foundation-sponsored research at the University of Milano (UniMi) has found that finasteride, a 5-alpha reductase (5α-R) inhibitor, also inhibits phenylethanolamine N-methyltransferase (PNMT), an enzyme found primarily in the adrenal gland, which in turn may play a role in producing some of the condition's most common side effects. (pfsfoundation.org)
- To better understand the mechanisms underlying the impact of stress on brain development and neuroendocrine function, this study focuses on the phenylethanolamine-N-methyltransferase (Pnmt) enzyme as a key mediator of stress hormone signaling. (ucf.edu)
Epinephrine5
- Phenylethanolamine N-methyltransferase (Noradrenaline N-Methyltransferase) catalyzes the conversion of norepinephrine (noradrenaline) to epinephrine (adrenaline). (proteopedia.org)
- And Phenylethanolamine n-methyltransferase converts norepinephrine into epinephrine (adrenaline). (nootropicsexpert.com)
- Tyrosine hydroxylase converts tyrosine to 3,4-dihydroxyphenylalanine (DOPA), which is converted to dopamine under the effect of aromatic l-amino acid decarboxylase, then dopamine β-hydroxylase converts dopamine to norepinephrine and phenylethanolamine N -methyltransferase converts norepinephrine to epinephrine. (medscape.com)
- Cell-free extracts from human full thickness skin (i.e., epidermis and dermis), suction blister roofs (i.e., epidermis) and from human keratinocytes express biopterin-dependent tyrosine hydroxylase a well as phenylethanolamine-N-methyl transferase, both representing key enzymes for the biosynthesis of epinephrine. (northwestern.edu)
- Catechol- O -methyltransferase is involved in the inactivation of the catecholamine neurotransmitters ( dopamine , epinephrine , and norepinephrine ). (en-academic.com)
MRNA1
- Furthermore, the mRNA levels of Th, Dbh, phenylethanolamine-N-methyltransferase, and P450scc were elevated by the HPG treatment when compared to the stress group. (korea.ac.kr)
Methionine1
- As part of this regulation, homocysteine is re-methylated to methionine via two different routes requiring either methionine synthase or betaine-homocysteine methyltransferase. (stackexchange.com)
Protein1
- In humans, catechol- O -methyltransferase protein is encoded by the COMT gene . (en-academic.com)
Catecholamine1
- Methyltransferases are very common in the catecholamine synthesis and deactivation pathways. (wikipedia.org)
Properties1
- It shares many properties found among the other methyltransferases. (wikipedia.org)
Levels1
- 2009) Glycine N -methyltransferase and regulation of S -adenosylmethionine levels. (stackexchange.com)
Gene2
Catalyzes1
- A methyltransferase that catalyzes the reaction of S-adenosyl-L-methionine and phenylethanolamine to yield S-adenosyl-L-homocysteine and N-methylphenylethanolamine. (bvsalud.org)
Found2
- It shares many properties found among the other methyltransferases. (wikipedia.org)
- Phenylethanolamine‐N‐methyltransferase, the enzymatic marker found only in adrenergic neurons, was detected immunocytochemically in terminals distributed throughout the area postrema and in neuronal perikarya and varicosities within the adjacent mNTS. (northwestern.edu)