Pyruvate Synthase
Pyruvate Kinase
Pyruvate Dehydrogenase Complex
A single eubacterial origin of eukaryotic pyruvate: ferredoxin oxidoreductase genes: implications for the evolution of anaerobic eukaryotes. (1/85)
The iron sulfur protein pyruvate: ferredoxin oxidoreductase (PFO) is central to energy metabolism in amitochondriate eukaryotes, including those with hydrogenosomes. Thus, revealing the evolutionary history of PFO is critical to understanding the origin(s) of eukaryote anaerobic energy metabolism. We determined a complete PFO sequence for Spironucleus barkhanus, a large fragment of a PFO sequence from Clostridium pasteurianum, and a fragment of a new PFO from Giardia lamblia. Phylogenetic analyses of eubacterial and eukaryotic PFO genes suggest a complex history for PFO, including possible gene duplications and horizontal transfers among eubacteria. Our analyses favor a common origin for eukaryotic cytosolic and hydrogenosomal PFOs from a single eubacterial source, rather than from separate horizontal transfers as previously suggested. However, with the present sampling of genes and species, we were unable to infer a specific eubacterial sister group for eukaryotic PFO. Thus, we find no direct support for the published hypothesis that the donor of eukaryote PFO was the common alpha-proteobacterial ancestor of mitochondria and hydrogenosomes. We also report that several fungi and protists encode proteins with PFO domains that are likely monophyletic with PFOs from anaerobic protists. In Saccharomyces cerevisiae, PFO domains combine with fragments of other redox proteins to form fusion proteins which participate in methionine biosynthesis. Our results are consistent with the view that PFO, an enzyme previously considered to be specific to energy metabolism in amitochondriate protists, was present in the common ancestor of contemporary eukaryotes and was retained, wholly or in part, during the evolution of oxygen-dependent and mitochondrion-bearing lineages. (+info)Structural and kinetic studies of the pyruvate-ferredoxin oxidoreductase/ferredoxin complex from Desulfovibrio africanus. (2/85)
The pyruvate-ferredoxin oxidoreductase (PFOR)/ferredoxin (Fd) system of Desulfovibrio africanus has been investigated with the aim of understanding more fully protein-protein interaction and the kinetic characteristics of electron transfer between the two redox partners. D. africanus contains three Fds (Fd I, Fd II and Fd III) able to function as electron acceptors for PFOR. The complete amino acid sequence of Fd II was determined by automatic Edman degradation. It revealed a striking similarity to that of Fd I. The protein consists of 64 residues and its amino acid sequence is in agreement with a molecular mass of 6822.5 Da as measured by electrospray MS. Fd II contains five cysteine residues of which the first four (Cys11, Cys14, Cys17 and Cys54) are likely ligands for the single [4Fe-4S] cluster. A covalently cross-linked complex between PFOR and Fd I or Fd II was obtained by using a water soluble carbodiimide. This complex exhibited a stoichiometry of one ferredoxin for one PFOR subunit and is dependent on the ionic strength. The second-order rate constants for electron transfer between PFOR and Fds determined electrochemically using cyclic voltammetry are 7 x 107 M-1.s-1 for Fd I and 2 x 107 M-1.s-1 for Fd II and Fd III. The Km values of PFOR for Fd I and Fd II measured both by the electrochemical and the spectrophotometric method have been found to be 3 microM and 5 microM, respectively. The three-dimensional modelling of Fd II and surface analysis of Fd I, Fd II and PFOR suggest that a protein-protein complex is likely to be formed between aspartic acid/glutamic acid invariant residues of Fds and lysine residues surrounding the distal [4Fe-4S] cluster of PFOR. All of these studies are indicative of the involvement of electrostatic interactions between the two redox partners. (+info)Rubredoxin from the green sulfur bacterium Chlorobium tepidum functions as an electron acceptor for pyruvate ferredoxin oxidoreductase. (3/85)
Rubredoxin (Rd) from the moderately thermophilic green sulfur bacterium Chlorobium tepidum was found to function as an electron acceptor for pyruvate ferredoxin oxidoreductase (PFOR). This enzyme, which catalyzes the conversion of pyruvate to acetyl-CoA and CO(2), exhibited an absolute dependence upon the presence of Rd. However, Rd was incapable of participating in the pyruvate synthase or CO(2) fixation reaction of C. tepidum PFOR, for which two different reduced ferredoxins are employed as electron donors. These results suggest a specific functional role for Rd in pyruvate oxidation and provide the initial indication that the two important physiological reactions catalyzed by PFOR/pyruvate synthase are dependent on different electron carriers in the cell. The UV-visible spectrum of oxidized Rd, with a monomer molecular weight of 6500, gave a molar absorption coefficient at 492 nm of 6.89 mM(-1) cm(-1) with an A(492)/A(280) ratio of 0.343 and contained one iron atom/molecule. Further spectroscopic studies indicated that the CD spectrum of oxidized C. tepidum Rd exhibited a unique absorption maximum at 385 nm and a shoulder at 420 nm. The EPR spectrum of oxidized Rd also exhibited unusual anisotropic resonances at g = 9.675 and g = 4.322, which is composed of a narrow central feature with broader shoulders to high and low field. The midpoint reduction potential of C. tepidum Rd was determined to be -87 mV, which is the most electronegative value reported for Rd from any source. (+info)The role of pyruvate ferredoxin oxidoreductase in pyruvate synthesis during autotrophic growth by the Wood-Ljungdahl pathway. (4/85)
Pyruvate:ferredoxin oxidoreductase (PFOR) catalyzes the oxidative decarboxylation of pyruvate to acetyl-CoA and CO(2). The catalytic proficiency of this enzyme for the reverse reaction, pyruvate synthase, is poorly understood. Conversion of acetyl-CoA to pyruvate links the Wood-Ljungdahl pathway of autotrophic CO(2) fixation to the reductive tricarboxylic acid cycle, which in these autotrophic anaerobes is the stage for biosynthesis of all cellular macromolecules. The results described here demonstrate that the Clostridium thermoaceticum PFOR is a highly efficient pyruvate synthase. The Michaelis-Menten parameters for pyruvate synthesis by PFOR are: V(max) = 1.6 unit/mg (k(cat) = 3.2 s(-1)), K(m)(Acetyl-CoA) = 9 micrometer, and K(m)(CO(2)) = 2 mm. The intracellular concentrations of acetyl-CoA, CoASH, and pyruvate have been measured. The predicted rate of pyruvate synthesis at physiological concentrations of substrates clearly is sufficient to support the role of PFOR as a pyruvate synthase in vivo. Measurements of its k(cat)/K(m) values demonstrate that ferredoxin is a highly efficient electron carrier in both the oxidative and reductive reactions. On the other hand, rubredoxin is a poor substitute in the oxidative direction and is inept in donating electrons for pyruvate synthesis. (+info)Comparison of sequencing of the por gene and typing of the opa gene for discrimination of Neisseria gonorrhoeae strains from sexual contacts. (5/85)
Typing of gonococcal strains is a valuable tool for the biological confirmation of sexual contacts. We have developed a typing method based on DNA sequencing of two overlapping por gene fragments generated by a heminested PCR. We compared sequencing of the por gene (POR sequencing) and typing of the opa gene (OPA typing) for the characterization of strains from 17 sexual partnerships. Both methods were highly discriminatory. A different genotype was detected in 15 of the 17 epidemiologically unconnected couples by POR sequencing and in 16 of the 17 couples by OPA typing with restriction enzyme HpaII. Within partnerships, identical genotypes were obtained from 16 of the 17 known sex contacts by POR sequencing and from 15 of the 17 by OPA typing. Compared to OPA typing, which relies on interpretation of bands in a gel, DNA sequence data offer the advantage of being objective and portable. As costs for sequencing decline, the method should become affordable for most laboratory personnel who wish to type gonococcal strains. (+info)Iron-induced changes in pyruvate metabolism of Tritrichomonas foetus and involvement of iron in expression of hydrogenosomal proteins. (6/85)
The main function of the hydrogenosome, a typical organelle of trichomonads, is to convert malate or pyruvate to H(2), CO(2) and acetate by a pathway associated with ATP synthesis. This pathway relies on activity of iron-sulfur proteins such as pyruvate:ferredoxin oxidoreductase (PFOR), hydrogenase and ferredoxin. To examine the effect of iron availability on proper hydrogenosomal function, the metabolic activity of the hydrogenosome and expression of hydrogenosomal enzymes were compared in Tritrichomonas foetus maintained under iron-rich (150 microM iron nitrilotriacetate) or iron-restricted (180 microM 2,2-dipyridyl) conditions in vitro. The activities of PFOR and hydrogenase, and also production of acetate and H(2), were markedly decreased or absent in iron-restricted trichomonads. Moreover, a decrease in activity of the hydrogenosomal malic enzyme, which is a non-Fe-S protein, was also observed. Impaired function of hydrogenosomes under iron-restricted conditions was compensated for by activation of the cytosolic pathway, mediating conversion of pyruvate to ethanol via acetaldehyde. This metabolic switch was fully reversible. Production of hydrogen by iron-restricted trichomonads was restored to the level of organisms grown under iron-rich conditions within 3 h after addition of 150 microM iron nitrilotriacetate. Protein analysis of purified hydrogenosomes from iron-restricted cells showed decreased levels of proteins corresponding to PFOR, malic enzyme and ferredoxin. Accordingly, these cells displayed decreased steady-state level and synthesis of mRNAs encoding PFOR and hydrogenosomal malic enzyme. These data demonstrate that iron is essential for function of the hydrogenosome, show its involvement in the expression of hydrogenosomal proteins and indicate the presence of iron-dependent control of gene transcription in Tt. foetus. (+info)Pyruvate : NADP+ oxidoreductase from the mitochondrion of Euglena gracilis and from the apicomplexan Cryptosporidium parvum: a biochemical relic linking pyruvate metabolism in mitochondriate and amitochondriate protists. (7/85)
Most eukaryotes perform the oxidative decarboxylation of pyruvate in mitochondria using pyruvate dehydrogenase (PDH). Eukaryotes that lack mitochondria also lack PDH, using instead the O(2)-sensitive enzyme pyruvate : ferredoxin oxidoreductase (PFO), which is localized either in the cytosol or in hydrogenosomes. The facultatively anaerobic mitochondria of the photosynthetic protist Euglena gracilis constitute a hitherto unique exception in that these mitochondria oxidize pyruvate with the O(2)-sensitive enzyme pyruvate : NADP oxidoreductase (PNO). Cloning and analysis of Euglena PNO revealed that the cDNA encodes a mitochondrial transit peptide followed by an N-terminal PFO domain that is fused to a C-terminal NADPH-cytochrome P450 reductase (CPR) domain. Two independent 5.8-kb full-size cDNAs for Euglena mitochondrial PNO were isolated; the gene was expressed in cultures supplied with 2% CO(2) in air and with 2% CO(2) in N(2). The apicomplexan Cryptosporidium parvum was also shown to encode and express the same PFO-CPR fusion, except that, unlike E. gracilis, no mitochondrial transit peptide for C. parvum PNO was found. Recombination-derived remnants of PNO are conserved in the genomes of Saccharomyces cerevisiae and Schizosaccharomyces pombe as proteins involved in sulfite reduction. Notably, Trypanosoma brucei was found to encode homologs of both PFO and all four PDH subunits. Gene organization and phylogeny revealed that eukaryotic nuclear genes for mitochondrial, hydrogenosomal, and cytosolic PFO trace to a single eubacterial acquisition. These findings suggest a common ancestry of PFO in amitochondriate protists with Euglena mitochondrial PNO and Cryptosporidium PNO. They are also consistent with the view that eukaryotic PFO domains are biochemical relics inherited from a facultatively anaerobic, eubacterial ancestor of mitochondria and hydrogenosomes. (+info)Spectroscopic analysis of desiccation-induced alterations of the chlorophyllide transformation pathway in etiolated barley leaves. (8/85)
Effects of water deficit on the chlorophyllide (Chlide) transformation pathway were studied in etiolated barley (Hordeum vulgare) leaves by analyzing absorption spectra and 77-K fluorescence spectra deconvoluted in components. Chlide transformations were examined in dehydrated leaves exposed to a 35-ms saturating flash triggering protochlorophyllide (Pchlide) and Chlide transformation processes. During the 90 min following the flash, we found that dehydration induced modifications of Chlide transformations, but no effect on Pchlide phototransformation into Chlide was observed. During this time, content of NADPH-Pchlide oxydoreductase in leaves did not change. Chlide transformation process in dehydrated leaves was characterized by the alteration of the Shibata shift process, by the appearance of a new Chlide species emitting at 692 nm, and by the favored formation of Chl(ide) A(668)F(676). The formation of Chl(ide) A(668)F(676), so-called "free Chlide," was probably induced by disaggregation of highly aggregated Chlide complexes. Here, we offer evidence for the alteration of photoactive Pchlide regeneration process, which may be caused by the desiccation-induced inhibition of Pchlide synthesis. (+info)I believe you may have meant to ask for the definition of "pyruvate dehydrogenase complex" rather than "pyruvate synthase," as I couldn't find any relevant medical information regarding a specific enzyme named "pyruvate synthase."
Pyruvate dehydrogenase complex (PDC) is a crucial enzyme complex in the human body, playing an essential role in cellular energy production. PDC is located within the mitochondrial matrix and catalyzes the oxidative decarboxylation of pyruvate, the end product of glycolysis, into acetyl-CoA. This process connects the glycolytic pathway to the citric acid cycle (Krebs cycle) and enables the continuation of aerobic respiration for efficient energy production in the form of ATP.
The pyruvate dehydrogenase complex consists of three main enzymes: pyruvate dehydrogenase (E1), dihydrolipoyl transacetylase (E2), and dihydrolipoyl dehydrogenase (E3). Additionally, two accessory proteins, E3-binding protein (E3BP) and protein X, are part of the complex. These enzymes work together to facilitate the conversion of pyruvate into acetyl-CoA, CO2, and NADH. Dysfunction in the pyruvate dehydrogenase complex can lead to various metabolic disorders and neurological symptoms.
Pyruvate kinase is an enzyme that plays a crucial role in the final step of glycolysis, a process by which glucose is broken down to produce energy in the form of ATP (adenosine triphosphate). Specifically, pyruvate kinase catalyzes the transfer of a phosphate group from phosphoenolpyruvate (PEP) to adenosine diphosphate (ADP), resulting in the formation of pyruvate and ATP.
There are several isoforms of pyruvate kinase found in different tissues, including the liver, muscle, and brain. The type found in red blood cells is known as PK-RBC or PK-M2. Deficiencies in pyruvate kinase can lead to a genetic disorder called pyruvate kinase deficiency, which can result in hemolytic anemia due to the premature destruction of red blood cells.
The Pyruvate Dehydrogenase Complex (PDC) is a multi-enzyme complex that plays a crucial role in cellular energy metabolism. It is located in the mitochondrial matrix and catalyzes the oxidative decarboxylation of pyruvate, the end product of glycolysis, into acetyl-CoA. This reaction links the carbohydrate metabolism (glycolysis) to the citric acid cycle (Krebs cycle), enabling the continuation of energy production in the form of ATP through oxidative phosphorylation.
The Pyruvate Dehydrogenase Complex consists of three main enzymes: pyruvate dehydrogenase (E1), dihydrolipoyl transacetylase (E2), and dihydrolipoyl dehydrogenase (E3). Additionally, two regulatory enzymes are associated with the complex: pyruvate dehydrogenase kinase (PDK) and pyruvate dehydrogenase phosphatase (PDP). These regulatory enzymes control the activity of the PDC through reversible phosphorylation and dephosphorylation, allowing the cell to adapt to varying energy demands and substrate availability.
Deficiencies or dysfunctions in the Pyruvate Dehydrogenase Complex can lead to various metabolic disorders, such as pyruvate dehydrogenase deficiency, which may result in neurological impairments and lactic acidosis due to disrupted energy metabolism.
 Pyruvate synthase
Pyruvate synthase
Pyruvate oxidase (CoA-acetylating)
Acetate-CoA ligase (ADP-forming)
Indolepyruvate ferredoxin oxidoreductase
Mitochondrial matrix
Unique properties of hyperthermophilic archaea
Thorarchaeota
N-acetylneuraminate synthase
Amino acid synthesis
1-deoxy-D-xylulose-5-phosphate synthase
PCK2
Glucose-1,6-bisphosphate synthase
1D-chiro-Inositol
Biosynthesis
Pyruvate, water dikinase
Thermales
Phenylacetylcarbinol
E1
Citramalate lyase
Ferredoxin
List of EC numbers (EC 1)
Halobacterium noricense
Glucagon
Deinococcus marmoris
Anthranilate synthase
2-ethylmalate synthase
Edward I. Solomon
Indole-3-glycerol-phosphate synthase
Fructolysis
Homocitrate synthase
Pyruvate synthase - Wikipedia
 NPL Proteins
NPL Proteins
 RCSB PDB - 1G6T: STRUCTURE OF EPSP SYNTHASE LIGANDED WITH SHIKIMATE-3-PHOSPHATE
RCSB PDB - 1G6T: STRUCTURE OF EPSP SYNTHASE LIGANDED WITH SHIKIMATE-3-PHOSPHATE
 SCOPe 2.04: Structural Classification of Proteins - extended
SCOPe 2.04: Structural Classification of Proteins - extended
 Citric acid cycle - Citizendium
Citric acid cycle - Citizendium
 Frontiers | Genetic Transformation of Sugarcane, Current Status and Future Prospects
Frontiers | Genetic Transformation of Sugarcane, Current Status and Future Prospects
Frontiers | Aluminum, a Friend or Foe of Higher Plants in Acid Soils
 Genes | Free Full-Text | Impacts of the Type I Toxin-Antitoxin System, SprG1/SprF1, on Staphylococcus aureus Gene Expression
Genes | Free Full-Text | Impacts of the Type I Toxin-Antitoxin System, SprG1/SprF1, on Staphylococcus aureus Gene Expression
 Michael Johnson | Department of Chemistry
Michael Johnson | Department of Chemistry
 CDD Conserved Protein Domain Family: dxs
CDD Conserved Protein Domain Family: dxs
 PDBeFold
PDBeFold
A High-Fat Diet Coordinately Downregulates Genes Required for Mitochondrial Oxidative Phosphorylation in Skeletal Muscle |...
 US Patent Application for Combination treatment for diabetes mellitus Patent Application (Application #20100179131 issued July...
US Patent Application for Combination treatment for diabetes mellitus Patent Application (Application #20100179131 issued July...
RCSB PDB - 6YAK: Split gene transketolase, active alpha2beta2 heterotetramer
 ID 2.7.9.2
ID 2.7.9.2
Molecular and Functional Characterization of theRhodopseudomonas palustris No. 7 Phosphoenolpyruvate Carboxykinase Gene |...
Bio2Vec
 Metapangenomics reveals depth-dependent shifts in metabolic potential for the ubiquitous marine bacterial SAR324 lineage |...
Metapangenomics reveals depth-dependent shifts in metabolic potential for the ubiquitous marine bacterial SAR324 lineage |...
 Diacetyl Formation by Oenococcus oeni during Winemaking Induced by Exogenous Pyruvate | American Journal of Enology and...
Diacetyl Formation by Oenococcus oeni during Winemaking Induced by Exogenous Pyruvate | American Journal of Enology and...
TCDB » SEARCH
Metabolic Flux-Based Modularity using Shortest Retroactive distances | BMC Systems Biology | Full Text
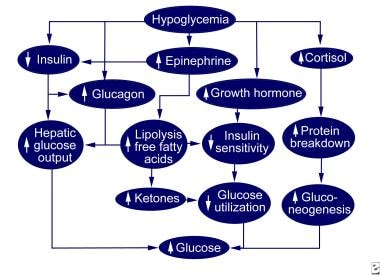
 Pediatric Hypoglycemia: Practice Essentials, Etiology, Epidemiology
Pediatric Hypoglycemia: Practice Essentials, Etiology, Epidemiology
 54-3716-1 - RIKEN Arabidopsis Genome Encyclopedia
54-3716-1 - RIKEN Arabidopsis Genome Encyclopedia
 SAUSA300 RS03745 - AureoWiki
SAUSA300 RS03745 - AureoWiki
Salivary gland proteome analysis of developing adult female Haemaphysalis longicornis ticks: molecular motor and TCA cycle...
 Targeting Metabolism to Induce Cell Death in Cancer Cells and Cancer Stem Cells
Targeting Metabolism to Induce Cell Death in Cancer Cells and Cancer Stem Cells
 Comparative Proteomic Analysis Reveals Novel Insights into the Interaction between Rice and Xanthomonas oryzae pv. oryzae |...
Comparative Proteomic Analysis Reveals Novel Insights into the Interaction between Rice and Xanthomonas oryzae pv. oryzae |...
Kinetic characterization of annotated glycolytic enzymes present in cellulose-fermenting Clostridium thermocellum suggests...
Citrate synthase6
- Also the enzymes citrate synthase, isocitrate dehydrogenase and alpha-ketoglutarate dehydrogenase, that regulate the first three steps of the TCA cycle, are inhibited by high concentrations of ATP. (citizendium.org)
- This includes both the entry point enzymes pyruvate dehydrogenase and citrate synthase. (citizendium.org)
- In contrast, acclimation to elevated temperatures had no effect on citrate synthase (CS) or pyruvate kinase (PK) activity in any tissue, nor were renal NKA or glutamine synthetase (GS) activities impacted. (mcmaster.ca)
- RESULTS: The levels of all enzymes were higher in the high-grade glioma group but only pyruvate dehydrogenase, citrate synthase, and isocitrate dehydrogenase levels showed statistical significance. (bvsalud.org)
- Acetyl dihydrolipoic acid is a substrate for citrate synthase, acetyl coenzyme A carboxylase and fatty acid synthetase. (ias.ac.in)
- for citrate synthase with acetyl dihydrolipoic acid was identical to acetyl coenzyme A (approximately 1 μmol citrate formed/min/mg protein) while the apparent K m was approximately 4 times higher with acetyl dihydrolipoic acid as the substrate. (ias.ac.in)
Dehydrogenase complex2
- Such enzymes include the pyruvate dehydrogenase complex that synthesises the acetyl-CoA needed for the first reaction of the TCA cycle. (citizendium.org)
- the E1 component of the pyruvate dehydrogenase complex and D-xylulose-5-phosphate synthase, in an attempt to rationalize differences in structure and substrate specificity between these enzymes. (rcsb.org)
Enzymes4
- In Escherichia coli , besides the PEPCK reaction, there is another pathway to form PEP from C 4 -dicarboxylic acid which involves the NAD- and NADP-dependent malic enzymes (MAEA and MAEB) and phosphoenolpyruvate synthase (PPS). (asm.org)
- The α-subunits of the oxaloacetate and methylmalonyl-CoA decarboxylases are homologous to many biotin-containing enzymes including (1) pyruvate carboxylases, (2) homocitrate synthases, (3) biotin carboxyl carrier proteins, (4) isopropylmalate synthases and (5) acyl-CoA carboxylase. (tcdb.org)
- By stimulating the expression of glucose transporters and glycolytic enzymes, HIF-1 promotes glycolysis to generate more pyruvate [ 5 ]. (hindawi.com)
- The main generation of H 2 S in cutaneous tissue mostly depends on enzymatic routes using L-cysteine and homocysteine by two pyridoxal-5 - phosphate-dependent enzymes, cystathionine β -synthase (CBS) and cystathionine γ -lyase (CSE). (hindawi.com)
Substrate4
- Glyphosate appears to occupy the binding site of the second substrate of EPSP synthase (phosphoenol pyruvate), mimicking an intermediate state of the ternary enzyme.substrates complex. (rcsb.org)
- Therefore, any substrate that increases intracellular pyruvate concentration can induce diacetyl accumulation. (ajevonline.org)
- The obtained crystal structure of acetohydroxyacid synthase AHAS, EC 2.2.1.6)in complex with herbicides of the sulphonylurea and imidazolinone family reveals the molecular basis for substrate/inhibitor binding. (riken.jp)
- Pyruvate is an important substrate in carbohydrate metabolism. (msdmanuals.com)
Subunit1
- DXP synthase is a thiamine diphosphate-dependent enzyme related to transketolase and the pyruvate dehydrogenase E1-beta subunit. (nih.gov)
Catalysed by pyruvate dehydrogenase1
- In aerobic organisms, this conversion is catalysed by pyruvate dehydrogenase, also uses thiamine pyrophosphate (TPP) but relies on lipoate as the electron acceptor. (wikipedia.org)
Metabolism4
- This enzyme participates in 4 metabolic pathways: pyruvate metabolism, propanoate metabolism, butanoate metabolism, and reductive carboxylate cycle (CO2 fixation). (wikipedia.org)
- In contrast to normal cells, most cancer cells predominantly produce energy by a high rate of glycolysis followed by lactate fermentation, even in the presence of oxygen, a less efficient metabolism compared to a low rate of glycolysis followed by mitochondrial oxidation of pyruvate [ 2 ]. (hindawi.com)
- Furthermore, HIF-1 actively limits the mitochondrial consumption of pyruvate at two levels: (i) through the enzyme PDK (PDH-kinase), which in turn inhibits PDH activity preventing the conversion of pyruvate into acetyl CoA, and thereby limiting mitochondrial metabolism of pyruvate [ 6 ] and (ii) through the direct activation of LDH [ 7 , 8 ]. (hindawi.com)
- Pyruvate metabolism. (msdmanuals.com)
Thiamine1
- By an acyloin condensation of pyruvate with glyceraldehyde 3-phosphate, it produces 1-deoxy-D-xylulose 5-phosphate, a precursor of thiamine diphosphate (TPP), pyridoxal phosphate, and the isoprenoid building block isopentenyl diphosphate (IPP). (nih.gov)
Dihydrodipicolinate Synthase3
- Find N-Acetylneuraminate Pyruvate Lyase (Dihydrodipicolinate Synthase) Proteins with a specific Grade. (antibodies-online.com)
- Find N-Acetylneuraminate Pyruvate Lyase (Dihydrodipicolinate Synthase) Proteins for a variety of species such as anti-Human N-Acetylneuraminate Pyruvate Lyase (Dihydrodipicolinate Synthase), anti-Xenopus laevis N-Acetylneuraminate Pyruvate Lyase (Dihydrodipicolinate Synthase), anti-Cow N-Acetylneuraminate Pyruvate Lyase (Dihydrodipicolinate Synthase). (antibodies-online.com)
- Find N-Acetylneuraminate Pyruvate Lyase (Dihydrodipicolinate Synthase) Proteins validated for a specific application such as ELISA, WB, SDS, AP. (antibodies-online.com)
Phosphoenolpyruvate synthase1
- AN phosphoenolpyruvate synthase. (expasy.org)
Decarboxylase2
Oxidation of pyruvate1
- The process begins with the oxidation of pyruvate, producing one CO 2 , and one acetyl-CoA. (citizendium.org)
Acetolactate2
- Furthermore, when additional pyruvate was added, α-acetolactate synthase ( alsS ) gene expression increased 1.6-fold and the diacetyl concentration increased from 0.4 mg/L to 2.3 mg/L. Although the highest alsS expression (a 10-fold increase) was found 24 hr after pyruvate addition, no further increase in diacetyl concentration was found. (ajevonline.org)
- Catalyzes the formation of acetolactate from pyruvate, the first step in valine and isoleucine biosynthesis. (riken.jp)
Biosynthesis1
- Biosynthesis of aromatic amino acids in plants, many bacteria, and microbes relies on the enzyme 5-enolpyruvylshikimate 3-phosphate (EPSP) synthase, a prime target for drugs and herbicides. (rcsb.org)
HRAC Group1
- Multiple resistance has evolved to herbicides in the Groups 2 (Legacy B), and Inhibition of Enolpyruvyl Shikimate Phosphate Synthase HRAC Group 9 (Legacy G). These particular biotypes are known to have resistance to chlorimuron-ethyl, glyphosate, and imazethapyr and they may be cross-resistant to other herbicides in the Groups 2 (Legacy B), and Inhibition of Enolpyruvyl Shikimate Phosphate Synthase HRAC Group 9 (Legacy G). (weedscience.org)
Cysteine1
- CSE mainly converts cysteine into H 2 S, pyruvate, and ammonia. (hindawi.com)
Glycolysis5
- Glycolysis breaks glucose (a six-carbon-molecule) down into pyruvate (a three-carbon molecule). (citizendium.org)
- When oxygen is limited, as in muscles that have undergone prolonged exercise, pyruvate is not consumed in the TCA cycle but is rather converted into lactic acid by lactate dehydrogenase (LDH) in a process termed anaerobic glycolysis. (hindawi.com)
- In glycolysis, the 6-carbon glucose is connected to two 3-carbon pyruvate molecules, and then to the 2-carbon acetyl-CoA. (encyclopedia.com)
- The breakdown of pyruvate irreversibly funnels the products of glycolysis into the Krebs cycle. (encyclopedia.com)
- Thus, the transformation of pyruvate to acetyl-CoA is the link between the metabolic reactions of glycolysis and the Krebs cycle. (encyclopedia.com)
Glutamine1
- This leads to accumulation of alanine and glutamine (transamination products of pyruvate and glutamate, respectively) and, finally, of ammonia. (medscape.com)
Molecule1
- The two hydrogenatoms removed from the pyruvate molecule yield NADH, which subsequently gives up its electrons to the electron transport chain to form ATP and water. (encyclopedia.com)
Enzyme3
- In enzymology, a pyruvate synthase (EC 1.2.7.1) is an enzyme that catalyzes the interconversion of pyruvate and acetyl-CoA. (wikipedia.org)
- The relevant equilibrium catalysed by PFOR is: pyruvate + CoA + oxidized ferredoxin ⇌ {\displaystyle \rightleftharpoons } acetyl-CoA + CO2 + reduced ferredoxin The 3 substrates of this enzyme are pyruvate, CoA, and oxidized ferredoxin, whereas its 3 products are acetyl-CoA, CO2, and reduced ferredoxin. (wikipedia.org)
- The systematic name of this enzyme class is pyruvate:ferredoxin 2-oxidoreductase (CoA-acetylating). (wikipedia.org)
Metabolic1
- Pyruvate, which is at the crossroad between lactate production and OXPHOS, constitutes a key metabolic intermediate. (hindawi.com)
Oxidative1
- Increase in the lactate:pyruvate ratio distinguishes oxidative phosphorylation defects from other genetic causes of lactic acidosis. (msdmanuals.com)
Synthetase2
- Other names in common use include: pyruvate oxidoreductase, pyruvate synthetase, pyruvate:ferredoxin oxidoreductase, pyruvic-ferredoxin oxidoreductase. (wikipedia.org)
- The research team combined 3-hydroxybenzoic acid synthetase (Hyg5) derived from Streptomyces hygroscopicus, 3-hydroxybenzoic acid 6-hydroxylase (3HB6H) derived from Rhodococcus jostii RHA, and maleyl pyruvate synthase (MPS) from Rhodococcus sp. (window-to-japan.eu)
Mitochondria3
- In eukaryotes , pyruvate moves into the mitochondria . (citizendium.org)
- In the presence of oxygen, the pyruvate is directed into mitochondria to be converted into acetyl CoA by the pyruvate dehydrogenase (PDH) or into alanine by transamination. (hindawi.com)
- Inside the mitochondria, pyruvate is completely oxidized through the TCA cycle, feeding reductive equivalents to the electron transport chain. (hindawi.com)
Phosphate1
- CA ATP + H2O + pyruvate = AMP + 2 H(+) + phosphate + phosphoenolpyruvate. (expasy.org)
NADH1
- Complejo enzimático que cataliza la DESCARBOXILACIÓN oxidativa y la DESAMINACIÓN de la GLICINA en DIÓXIDO DE CARBONO, AMONÍACO, NADH y N5N10-metilenotetrahidrofolato. (bvsalud.org)
Acetyl3
- N-acetylneuraminate lyases regulate cellular concentrations of N-acetyl-neuraminic acid (sialic acid) by mediating the reversible conversion of sialic acid into N-acetylmannosamine and pyruvate. (antibodies-online.com)
- This regulation ensures that the TCA cycle will not oxidise excessive amounts of pyruvate and acetyl-CoA when ATP in the cell is plentiful. (citizendium.org)
- In an earlier paper the formation of acetyl dihydrolipoic from pyruvate and dihydrolipoic acid catalyzed by pyruvate dehydrogenase has been reported. (ias.ac.in)
Gene1
- Studies of a pckA-lacZ gene fusion indicated that when cells were grown in minimal media with various carbon sources, such as succinate, malate, pyruvate, lactate, or ethanol, under both anaerobic light and aerobic dark conditions, the pckA gene was induced in log phase, irrespective of the carbon source. (asm.org)
Metabolite1
- Tizoxanide, an active metabolite of nitazoxanide Amixicile, a water-soluble derivative of nitazoxanide, is a potent inhibitor of pyruvate:ferredoxin oxidoreductase and is in pre-clinical studies to treat infections of Helicobacter pylori and Clostridium difficile. (wikipedia.org)
Protein1
- Image analysis of the matched maps identified 7 proteins that were either over- or down-expressed: activated protein kinase c receptor (LACK), alpha tubulin (×2), prostaglandin f2-alpha synthase, protein disulfide isomerase, vesicular transport protein and a hypothetical protein. (who.int)
Herbicides1
- The elucidation of the active site of EPSP synthase and especially of the binding pattern of glyphosate provides a valuable roadmap for engineering new herbicides and herbicide-resistant crops, as well as new antibiotic and antiparasitic drugs. (rcsb.org)
Carbon1
- An PFOR reaction starts with the nucleophilic attack of C2 of TPP on the 2-oxo carbon of pyruvate, which forms a lactyl-TPP adduct. (wikipedia.org)
Central1
- Порушення метаболізму пірувату Inability to metabolize pyruvate causes lactic acidosis and a variety of central nervous system abnormalities. (msdmanuals.com)
Formation2
- This study evaluates the effect of exogenous pyruvate on diacetyl formation and the expression of diacetyl-related genes in Oenococcus oeni during winemaking. (ajevonline.org)
- Diacetyl formation by Oenococcus oeni was induced by yeast-derived pyruvate in the early stage of winemaking. (ajevonline.org)
Water2
- ID 2.7.9.2 DE pyruvate, water dikinase. (expasy.org)
- AN pyruvate,water dikinase. (expasy.org)
Normal3
- In normal cells, the fate of pyruvate depends on many factors, one of which is oxygen availability. (hindawi.com)
- Intraperitoneal administration of lipoic acid (10 mg/100 g) does not effect changes in serum insulin levels in normal and alloxan diabetic rats, while normalising increased serum pyruvate, and impaired liver pyruvic dehydrogenase characteristic of the diabetic state. (ias.ac.in)
- in which the lactate:pyruvate ratio remains normal. (msdmanuals.com)
Transport1
- L'analyse d'image des cartes appariées a permis d'identifier sept protéines qui étaient soit surexprimées, soit sous-exprimées : le récepteur de la protéine kinase C activée, la tubuline alpha (×2), la prostaglandine F2-alpha synthase, la protéine disulfide isomérase, une protéine du transport vésiculaire et une protéine hypothétique. (who.int)