Dihydropteridine Reductase
Pteridines
GTP Cyclohydrolase
Characterization of a novel unconjugated pteridine glycoside, cyanopterin, in Synechocystis sp. PCC 6803. (1/356)
A new pteridine glycoside, called cyanopterin, was isolated from Synechocystis sp. PCC 6803 and its structure was elucidated as 6-[1-(4-O-methyl-(alpha-d-glucuronyl)-(1, 6)-(beta-d-galactosyloxy]methylpterin by chemical degradation and 1H- and 13C-NMR spectroscopic means. Cyanopterin is constitutively synthesized at a relatively high intracellular concentration that is comparable to that of chlorophyll a in a molar ratio of approximately 1 to 1.6. The in vivo oxidation state of cyanopterin is primarily the fully reduced 5,6,7,8-tetrahydro form. The cellular function is unknown at present. The findings have established a model system, using Synechocystis sp. PCC 6803, for studies of the physiological functions of unconjugated pteridine glycosides found mostly in cyanobacteria. (+info)A corrinoid-dependent catabolic pathway for growth of a Methylobacterium strain with chloromethane. (2/356)
Methylobacterium sp. strain CM4, an aerobic methylotrophic alpha-proteobacterium, is able to grow with chloromethane as a carbon and energy source. Mutants of this strain that still grew with methanol, methylamine, or formate, but were unable to grow with chloromethane, were previously obtained by miniTn5 mutagenesis. The transposon insertion sites in six of these mutants mapped to two distinct DNA fragments. The sequences of these fragments, which extended over more than 17 kb, were determined. Sequence analysis, mutant properties, and measurements of enzyme activity in cell-free extracts allowed the definition of a multistep pathway for the conversion of chloromethane to formate. The methyl group of chloromethane is first transferred by the protein CmuA (cmu: chloromethane utilization) to a corrinoid protein, from where it is transferred to H4folate by CmuB. Both CmuA and CmuB display sequence similarity to methyltransferases of methanogenic archaea. In its C-terminal part, CmuA is also very similar to corrinoid-binding proteins, indicating that it is a bifunctional protein consisting of two domains that are expressed as separate polypeptides in methyl transfer systems of methanogens. The methyl group derived from chloromethane is then processed by means of pterine-linked intermediates to formate by a pathway that appears to be distinct from those already described in Methylobacterium. Remarkable features of this pathway for the catabolism of chloromethane thus include the involvement of a corrinoid-dependent methyltransferase system for dehalogenation in an aerobe and a set of enzymes specifically involved in funneling the C1 moiety derived from chloromethane into central metabolism. (+info)Enzymatic synthesis of biopterin from D-erythrodihydroneopterin triphosphate by extracts of kidneys from Syrian golden hamsters. (3/356)
An enzyme system was found in either crude homogenates of dialyzed extracts of liver, kidney, lung, and brain from Syrian golden hamsters that catalyzed the synthesis of radioactive 6(L-erythro-1',2'-dihydroxypropyl)pterin (biopterin) from [U-14C]6(D-erythro-1',2',3'-trihydroxypropyl)-7,8-dihydropterin triphosphate (D-erythrolH2neopterin-PPP) preparation. The specific radioactivity of biopterin was found to be comparable to that of D-erythroH2neopterin-PPP. The enzyme system from hamster kidney was purified severalfold by fractionation with ammonium sulfate and with an Ultrogel AcA-34 column. It was demonstrated that (a) NADPH or NADAH was essential and that (b) Mg2+ was stimulatory for the enzymatic synthesis of biopterin from D-erythroH2-NEOPTERIN-PPP. Also GTP and nonphosphorylated neopterins were not converted to biopterin. Although 6-lactyl-7,8-dihydropterin (sepiapterin) was converted to biopterin in the presence of NADPH, sepiapterin was not detected from D-erythroH2neopterin-PPP in the absence of NADPH. A preliminary experiment was performed to identify dihydrobiopterin. (+info)The energy conserving methyltetrahydromethanopterin:coenzyme M methyltransferase complex from methanogenic archaea: function of the subunit MtrH. (4/356)
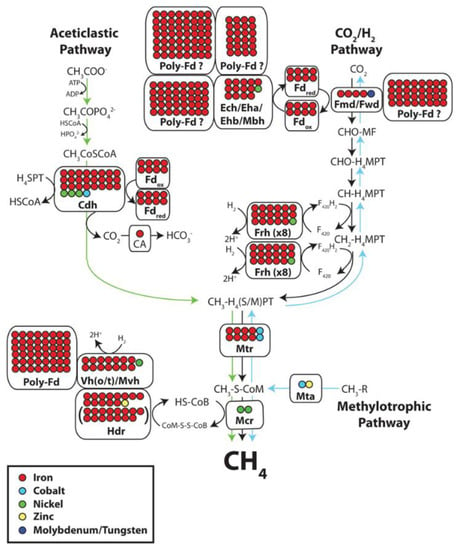
In methanogenic archaea the transfer of the methyl group of N5-methyltetrahydromethanopterin to coenzyme M is coupled with energy conservation. The reaction is catalyzed by a membrane associated multienzyme complex composed of eight different subunits MtrA-H. The 23 kDa subunit MtrA harbors a corrinoid prosthetic group which is methylated and demethylated in the catalytic cycle. We report here that the 34 kDa subunit MtrH catalyzes the methylation reaction. MtrH was purified and shown to exhibit methyltetrahydromethanopterin:cob(I)alamin methyltransferase activity. Sequence comparison revealed similarity of MtrH with MetH from Escherichia coli and AcsE from Clostridium thermoaceticum: both enzymes exhibit methyltetrahydrofolate:cob(I)alamin methyltransferase activity. (+info)Activation of neuronal nitric-oxide synthase by the 5-methyl analog of tetrahydrobiopterin. Functional evidence against reductive oxygen activation by the pterin cofactor. (5/356)
Tetrahydrobiopterin ((6R)-5,6,7,8-tetrahydro-L-biopterin (H4biopterin)) is an essential cofactor of nitric-oxide synthases (NOSs), but its role in enzyme function is not known. Binding of the pterin affects the electronic structure of the prosthetic heme group in the oxygenase domain and results in a pronounced stabilization of the active homodimeric structure of the protein. However, these allosteric effects are also produced by the potent pterin antagonist of NOS, 4-amino-H4biopterin, suggesting that the natural cofactor has an additional, as yet unknown catalytic function. Here we show that the 5-methyl analog of H4biopterin, which does not react with O2, is a functionally active pterin cofactor of neuronal NOS. Activation of the H4biopterin-free enzyme occurred in a biphasic manner with half-maximally effective concentrations of approximately 0.2 microM and 10 mM 5-methyl-H4biopterin. Thus, the affinity of the 5-methyl compound was 3 orders of magnitude lower than that of the natural cofactor, allowing the direct demonstration of the functional anticooperativity of the two pterin binding sites of dimeric NOS. In contrast to H4biopterin, which inactivates nitric oxide (NO) through nonenzymatic superoxide formation, up to 1 mM of the 5-methyl derivative did not consume O2 and had no effect on NO steady-state concentrations measured electrochemically with a Clark-type NO electrode. Therefore, reconstitution with 5-methyl-H4biopterin allowed, for the first time, the detection of enzymatic NO formation in the absence of superoxide or NO scavengers. These results unequivocally identify free NO as a NOS product and indicate that reductive O2 activation by the pterin cofactor is not essential to NO biosynthesis. (+info)Tyrosine hydroxylase binds tetrahydrobiopterin cofactor with negative cooperativity, as shown by kinetic analyses and surface plasmon resonance detection. (6/356)
Kinetic studies of tetrameric recombinant human tyrosine hydroxylase isoform 1 (hTH1) have revealed properties so far not reported for this enzyme. Firstly, with the natural cofactor (6R)-Lerythro-5,6,7, 8-tetrahydrobiopterin (H4biopterin) a time-dependent change (burst) in enzyme activity was observed, with a half-time of about 20 s for the kinetic transient. Secondly, nonhyperbolic saturation behaviour was found for H4biopterin with a pronounced negative cooperativity (0.39 < h < 0.58; [S]0.5 = 24 +/- 4 microM). On phosphorylation of Ser40 by protein kinase A, the affinity for H4biopterin increased ([S]0.5 = 11 +/- 2 microM) and the negative cooperativity was amplified (h = 0.27 +/- 0.03). The dimeric C-terminal deletion mutant (Delta473-528) of hTH1 also showed negative cooperativity of H4biopterin binding (h = 0.4). Cooperativity was not observed with the cofactor analogues 6-methyl-5,6,7,8-tetrahydropterin (h = 0.9 +/- 0.1; Km = 62.7 +/- 5.7 microM) and 3-methyl-5,6,7, 8-tetrahydropterin (H43-methyl-pterin)(h = 1.0 +/- 0.1; Km = 687 +/- 50 microM). In the presence of 1 mM H43-methyl-pterin, used as a competitive cofactor analogue to BH4, hyperbolic saturation curves were also found for H4biopterin (h = 1.0), thus confirming the genuine nature of the kinetic negative cooperativity. This cooperativity was confirmed by real-time biospecific interaction analysis by surface plasmon resonance detection. The equilibrium binding of H4biopterin to the immobilized iron-free apoenzyme results in a saturable positive resonance unit (DeltaRU) response with negative cooperativity (h = 0.52-0.56). Infrared spectroscopic studies revealed a reduced thermal stability both of the apo-and the holo-hTH1 on binding of H4biopterin and Lerythro-dihydrobiopterin (H2biopterin). Moreover, the ligand-bound forms of the enzyme also showed a decreased resistance to limited tryptic proteolysis. These findings indicate that the binding of H4biopterin at the active site induces a destabilizing conformational change in the enzyme which could be related to the observed negative cooperativity. Thus, our studies provide new insight into the regulation of TH by the concentration of H4biopterin which may have significant implications for the physiological regulation of catecholamine biosynthesis in neuroendocrine cells. (+info)2.0 A X-ray structure of the ternary complex of 7,8-dihydro-6-hydroxymethylpterinpyrophosphokinase from Escherichia coli with ATP and a substrate analogue. (7/356)
The X-ray crystal structure of 7,8-dihydro-6-hydroxymethylpterinpyrophosphokinase (PPPK) in a ternary complex with ATP and a pterin analogue has been solved to 2.0 A resolution, giving, for the first time, detailed information of the PPPK/ATP intermolecular interactions and the accompanying conformational change. The first 100 residues of the 158 residue peptide contain a betaalpha betabeta alphabeta motif present in several other proteins including nucleoside diphosphate kinase. Comparative sequence examination of a wide range of prokaryotic and lower eukaryotic species confirms the conservation of the PPPK active site, indicating the value of this de novo folate biosynthesis pathway enzyme as a potential target for the development of novel broad-spectrum anti-infective agents. (+info)Distribution of tetrahydromethanopterin-dependent enzymes in methylotrophic bacteria and phylogeny of methenyl tetrahydromethanopterin cyclohydrolases. (8/356)
The methylotrophic proteobacterium Methylobacterium extorquens AM1 possesses tetrahydromethanopterin (H(4)MPT)-dependent enzymes, which are otherwise specific to methanogenic and sulfate-reducing archaea and which have been suggested to be involved in formaldehyde oxidation to CO(2) in M. extorquens AM1. The distribution of H(4)MPT-dependent enzyme activities in cell extracts of methylotrophic bacteria from 13 different genera are reported. H(4)MPT-dependent activities were detected in all of the methylotrophic and methanotrophic proteobacteria tested that assimilate formaldehyde by the serine or ribulose monophosphate pathway. H(4)MPT-dependent activities were also found in autotrophic Xanthobacter strains. However, no H(4)MPT-dependent enzyme activities could be detected in other autotrophic alpha-proteobacteria or in gram-positive methylotrophic bacteria. Genes encoding methenyl H(4)MPT cyclohydrolase (mch genes) were cloned and sequenced from several proteobacteria. Bacterial and archaeal Mch sequences have roughly 35% amino acid identity and form distinct groups in phylogenetic analysis. (+info)Pterins are a group of naturally occurring pigments that are derived from purines. They are widely distributed in various organisms, including bacteria, fungi, and animals. In humans, pterins are primarily found in the eye, skin, and hair. Some pterins have been found to play important roles as cofactors in enzymatic reactions and as electron carriers in metabolic pathways.
Abnormal levels of certain pterins can be indicative of genetic disorders or other medical conditions. For example, an excess of biopterin, a type of pterin, is associated with phenylketonuria (PKU), a genetic disorder that affects the body's ability to metabolize the amino acid phenylalanine. Similarly, low levels of neopterin, another type of pterin, can be indicative of immune system dysfunction or certain types of cancer.
Medical professionals may measure pterin levels in blood, urine, or other bodily fluids to help diagnose and monitor these conditions.
Dihydropteridine reductase is an enzyme that plays a crucial role in the metabolism of certain amino acids, specifically phenylalanine and tyrosine. This enzyme is responsible for reducing dihydropteridines to tetrahydropteridines, which is a necessary step in the regeneration of tetrahydrobiopterin (BH4), an essential cofactor for the enzymes phenylalanine hydroxylase and tyrosine hydroxylase.
Phenylalanine hydroxylase and tyrosine hydroxylase are involved in the conversion of the amino acids phenylalanine and tyrosine to tyrosine and dopa, respectively. Without sufficient BH4, these enzymes cannot function properly, leading to an accumulation of phenylalanine and a decrease in the levels of important neurotransmitters such as dopamine, norepinephrine, and serotonin.
Deficiency in dihydropteridine reductase can lead to a rare genetic disorder known as dihydropteridine reductase deficiency (DPRD), which is characterized by elevated levels of phenylalanine and neurotransmitter imbalances, resulting in neurological symptoms such as developmental delay, seizures, and hypotonia. Treatment typically involves a low-phenylalanine diet and supplementation with BH4.
Pteridines are a class of heterocyclic aromatic organic compounds that are structurally related to pterins, which contain a pyrimidine ring fused to a pyrazine ring. They are naturally occurring substances that can be found in various living organisms such as bacteria, fungi, plants, and animals.
Pteridines have several important biological functions. For instance, they play a crucial role in the synthesis of folate and biotin, which are essential cofactors for various metabolic reactions in the body. Additionally, some pteridines function as chromophores, contributing to the coloration of certain organisms such as butterflies and birds.
In medicine, pteridines have been studied for their potential therapeutic applications. For example, some synthetic pteridine derivatives have shown promising results in preclinical studies as antitumor, antiviral, and antibacterial agents. However, further research is needed to fully understand the medical implications of these compounds.
Biopterin is a type of pteridine compound that acts as a cofactor in various biological reactions, particularly in the metabolism of amino acids such as phenylalanine and tyrosine. It plays a crucial role in the production of neurotransmitters like dopamine, serotonin, and noradrenaline. Biopterin exists in two major forms: tetrahydrobiopterin (BH4) and dihydrobiopterin (BH2). BH4 is the active form that participates in enzymatic reactions, while BH2 is an oxidized form that can be reduced back to BH4 by the action of dihydrobiopterin reductase.
Deficiencies in biopterin metabolism have been linked to several neurological disorders, including phenylketonuria (PKU), dopamine-responsive dystonia, and certain forms of autism. In these conditions, the impaired synthesis or recycling of biopterin can lead to reduced levels of neurotransmitters, causing various neurological symptoms.
GTP Cyclohydrolase is a crucial enzyme in the biosynthetic pathway of neurotransmitters and other biogenic amines. It catalyzes the conversion of GTP (guanosine triphosphate) to dihydroneopterin triphosphate, which is a key intermediate in the production of tetrahydrobiopterin (BH4). Tetrahydrobiopterin serves as a cofactor for various enzymes involved in the synthesis of neurotransmitters such as dopamine, serotonin, and noradrenaline.
There are two main isoforms of GTP Cyclohydrolase: GTPCH1 (GTP Cyclohydrolase 1) and GTPCH2 (GTP Cyclohydrolase 2). GTPCH1 is primarily expressed in the brain, kidneys, and lungs, while GTPCH2 is mainly found in the liver. Defects or mutations in the GTPCH1 gene can lead to a rare genetic disorder known as Dopa-Responsive Dystonia (DRD), which is characterized by symptoms such as muscle stiffness, involuntary movements, and Parkinsonism.
 Pterin - Wikipedia
Pterin - Wikipedia Whittakers IOM Pt Erin - Whittaker Trading Ltd
Whittakers IOM Pt Erin - Whittaker Trading Ltd Spring tree bare - Pt Erin park - Juliet Batten
Spring tree bare - Pt Erin park - Juliet Batten Three histidines in pterin-4a-carbinolamine dehydratase are the important residues for substrate binding and catalysis
Three histidines in pterin-4a-carbinolamine dehydratase are the important residues for substrate binding and catalysis MedlinePlus: Genes: P
MedlinePlus: Genes: P Influencing Factors on the Use of Tetrahydrobiopterin in Patients with Phenylketonuria
Influencing Factors on the Use of Tetrahydrobiopterin in Patients with Phenylketonuria PET Scanning in Autism Spectrum Disorder: Overview, Evaluation of Autism Spectrum Disorder, Screening for PET Scanning
PET Scanning in Autism Spectrum Disorder: Overview, Evaluation of Autism Spectrum Disorder, Screening for PET Scanning A fossilized snake shows its true colors | ScienceDaily
A fossilized snake shows its true colors | ScienceDaily Human ZNF3 cDNA Clone BC025265 pUC Plasmid | CSB-CL026651HU4 | Cusabio
Human ZNF3 cDNA Clone BC025265 pUC Plasmid | CSB-CL026651HU4 | Cusabio Antioxidants | Free Full-Text | Neopterin, Inflammation, and Oxidative Stress: What Could We Be Missing?
Antioxidants | Free Full-Text | Neopterin, Inflammation, and Oxidative Stress: What Could We Be Missing? Advanced Search Results - Public Health Image Library(PHIL)
Advanced Search Results - Public Health Image Library(PHIL) MBS9933280 | Rat Phosphatidylinositol 3,4,5-Trisphosphate-Dependent Rac Exchanger 1 Protein (PREX1) ELISA Kit | MyBiosource
MBS9933280 | Rat Phosphatidylinositol 3,4,5-Trisphosphate-Dependent Rac Exchanger 1 Protein (PREX1) ELISA Kit | MyBiosource Aminopterin
Summary Report | CureHunter
Aminopterin
Summary Report | CureHunter Thieme E-Journals - Journal of Pediatric Biochemistry / Abstract
Thieme E-Journals - Journal of Pediatric Biochemistry / Abstract Human Metabolome Database: Showing metabocard for Citicoline (HMDB0001413)
Human Metabolome Database: Showing metabocard for Citicoline (HMDB0001413) WikiGenes - Ranachrome 4 - 2-amino-1,8-dihydropteridine-4,7-dione
WikiGenes - Ranachrome 4 - 2-amino-1,8-dihydropteridine-4,7-dione Almut KELBER | Professor of Sensory Biology | Dr.rer.nat | Lund University, Lund | LU | Department of Biology | Research profile
Almut KELBER | Professor of Sensory Biology | Dr.rer.nat | Lund University, Lund | LU | Department of Biology | Research profile Richard E. Lee, PhD - St. Jude Children's Research Hospital
Richard E. Lee, PhD - St. Jude Children's Research Hospital Zeitschrift für Naturforschung B Volume 20 Issue 4
Zeitschrift für Naturforschung B Volume 20 Issue 4 SCOPe 2.08: Domain d1vaxd2: 1vax D:11-141
SCOPe 2.08: Domain d1vaxd2: 1vax D:11-141