Enteropeptidase
Trypsinogen
Purification and further characterization of enteropeptidase from porcine duodenum. (1/101)
Enteropeptidase [EC 3.4.21.9] is a membrane-bound serine endopeptidase present in the duodenum that converts trypsinogen to trypsin. We previously cloned the cDNA of the porcine enzyme and deduced its entire amino acid sequence [M. Matsushima et al. (1994) J. Biol. Chem. 269, 19976-19982]. In the present study, we purified the porcine enzyme approximately 2,200-fold in a 12% yield from a duodenal mucosal extract to apparent homogeneity by an improved procedure comprising four steps of chromatography including benzamidine-Sepharose affinity chromatography. Lectin blotting analysis suggested that the enzyme is glycosylated mainly with N-linked carbohydrate chains of the tri- and/or tetraantennary complex type. The H and L chains of the enzyme were separated into two major bands upon SDS-PAGE under reducing conditions, suggesting that the enzyme mainly comprises two isoforms, a higher molecular weight form and a lower molecular weight form. The enzyme was also separated by lectin affinity chromatography into two major fractions, named isoforms I and II, which corresponded to the higher and lower molecular weight forms, respectively. These two isoforms appeared to be different only in the carbohydrate moiety, having essentially the same enzymatic properties. The enzyme was optimally active at pH 8.0 toward Gly-Asp-Asp-Asp-Asp-Lys-beta-naphthylamide, and was inhibited strongly by various serine proteinase inhibitors. Furthermore, it was also strongly inhibited by E-64 [L-trans-epoxysuccinyl-leucylamide-(4-guanido)-butane], a cysteine proteinase inhibitor. Substrate specificity studies involving various synthetic peptides indicated that acidic residues at the P2, P3, and/or P4 positions are especially favorable for maximal activity, but are not absolutely necessary, at least in the cases of peptide substrates. (+info)Hereditary pancreatitis-associated mutation asn(21) --> ile stabilizes rat trypsinogen in vitro. (2/101)
Mutations Arg(117) --> His and Asn(21) --> Ile in human trypsinogen-I have been recently associated with hereditary pancreatitis (HP). The Arg(117) --> His substitution is believed to cause pancreatitis by stabilizing trypsin against autolytic degradation, while the mechanism of action of Asn(21) --> Ile has been unknown. In an effort to understand the effect(s) of this mutation, Thr(21) in the highly homologous rat trypsinogen-II was replaced with Asn or Ile, and the recombinant zymogens and their active trypsin forms were studied. Kinetic parameters of all three trypsins were comparable, and the active enzymes suffered autolysis at similar rates, indicating that neither catalytic properties nor proteolytic stability of trypsin are influenced by mutations at position 21. When incubated at pH 8.0, 37 degrees C, pure zymogens underwent autoactivation with concomitant trypsinolytic degradation in a Ca(2+)-dependent fashion. Thus, in the presence of 5 mM Ca(2+), autoactivation and digestion of the zymogens after Arg(117) and Lys(188) were observed, while in the presence of 1 mM EDTA autoactivation and cleavage at Lys(188) were reduced, and zymogenolysis at the Arg(117) site was enhanced. Overall rates of zymogen degradation in [Asn(21)]- and [Ile(21)]trypsinogens were higher in Ca(2+) than in EDTA, while [Thr(21)]trypsinogen demonstrated inverse characteristics. Remarkably, both in the presence and absence of Ca(2+), [Ile(21)]trypsinogen exhibited significantly higher stability against autoactivation and proteolysis than zymogens with Asn(21) or Thr(21). The observations suggest that autocatalytic trypsinogen degradation may be an important defense mechanism against excessive trypsin generation in the pancreas, and trypsinogen stabilization by the Asn(21) --> Ile mutation plays a role in the pathogenesis of HP. (+info)Trypsin and activation of circulating trypsinogen contribute to pancreatitis-associated lung injury. (3/101)
Pancreatic proteases are secreted in acute pancreatitis, but their contribution to associated lung injury is unclear. Applying models of mild edematous (intravenous caerulein) and severe necrotizing (intraductal glycodeoxycholic acid) pancreatitis in rats, we showed that both trypsinogen and trypsin concentrations in peripheral blood, as well as lung injury, correlate with the severity of the disease. To isolate the potential contribution of proteases to lung injury, trypsin or trypsinogen was injected into healthy rats or trypsinogen secreted in caerulein pancreatitis was activated by intravenous enterokinase. Pulmonary injury induced by protease infusions was dose dependent and was ameliorated by neutrophil depletion. Trypsinogen activation worsened lung injury in mild pancreatitis. In vitro incubation of leukocytes with trypsinogen showed that stimulated leukocytes can convert trypsinogen to trypsin. In conclusion, this study demonstrates that the occurrence and severity of pancreatitis-associated lung injury (PALI) corresponds to the levels of circulating trypsinogen and its activation to trypsin. Neutrophils are involved in both protease activation and development of pulmonary injury. (+info)Formation of enzymatically active, homotypic, and heterotypic tetramers of mouse mast cell tryptases. Dependence on a conserved Trp-rich domain on the surface. (4/101)
Mouse mast cell protease (mMCP) 6 and mMCP-7 are homologous tryptases stored in granules as macromolecular complexes with heparin and/or chondroitin sulfate E containing serglycin proteoglycans. When pro-mMCP-7 and pseudozymogen forms of this tryptase and mMCP-6 were separately expressed in insect cells, all three recombinant proteins were secreted into the conditioned medium as properly folded, enzymatically inactive 33-kDa monomers. However, when their propeptides were removed, mMCP-6 and mMCP-7 became enzymatically active and spontaneously assumed an approximately 150-kDa tetramer structure. Heparin was not required for this structural change. When incubated at 37 degrees C, recombinant mMCP-7 progressively lost its enzymatic activity in a time-dependent manner. Its N-linked glycans helped regulate the thermal stability of mMCP-7. However, the ability of this tryptase to form the enzymatically active tetramer was more dependent on a highly conserved Trp-rich domain on its surface. Although recombinant mMCP-6 and mMCP-7 preferred to form homotypic tetramers, these tryptases readily formed heterotypic tetramers in vitro. This latter finding indicates that the tetramer structural unit is a novel way the mast cell uses to assemble varied combinations of tryptases. (+info)A protein disulfide isomerase gene fusion expression system that increases the extracellular productivity of Bacillus brevis. (5/101)
We have developed a versatile Bacillus brevis expression and secretion system based on the use of fungal protein disulfide isomerase (PDI) as a gene fusion partner. Fusion with PDI increased the extracellular production of heterologous proteins (light chain of immunoglobulin G, 8-fold; geranylgeranyl pyrophosphate synthase, 12-fold). Linkage to PDI prevented the aggregation of the secreted proteins, resulting in high-level accumulation of fusion proteins in soluble and biologically active forms. We also show that the disulfide isomerase activity of PDI in a fusion protein is responsible for the suppression of the aggregation of the protein with intradisulfide, whereas aggregation of the protein without intradisulfide was prevented even when the protein was fused to a mutant PDI whose two active sites were disrupted, suggesting that another PDI function, such as chaperone-like activity, synergistically prevented the aggregation of heterologous proteins in the PDI fusion expression system. (+info)Activation of recombinant proenteropeptidase by duodenase. (6/101)
Duodenase, a serine proteinase from bovine Brunner's (duodenal) glands that was predicted to be a natural activator of enteropeptidase zymogen, cleaves and activates recombinant single-chain bovine proenteropeptidase (kcat/Km = 2700 M(-1) s(-1)). The measured rate of proenteropeptidase cleavage by duodenase was about 70-fold lower compared with the rate of trypsin-mediated cleavage of the zymogen. The role of duodenase is supposed to be the primary activator of proenteropeptidase maintaining a certain level of active enteropeptidase in the duodenum. A new scheme of proteolytic activation cascade of digestive proteases is discussed. (+info)Human cationic trypsinogen. Role of Asn-21 in zymogen activation and implications in hereditary pancreatitis. (7/101)
Mutation Asn-21 --> Ile in human cationic trypsinogen (Tg-1) has been associated with hereditary pancreatitis. Recent studies with rat anionic Tg (Tg-2) indicated that the analogous Thr-21 --> Ile mutation stabilizes the zymogen against autoactivation, whereas it has no effect on catalytic properties or autolytic stability of trypsin (Sahin-Toth, M. (1999) J. Biol. Chem. 274, 29699-29704). In the present paper, human cationic Tg (Asn-21-Tg) and mutants Asn-21 --> Ile (Ile-21-Tg) and Asn-21 --> Thr (Thr-21-Tg) were expressed in Escherichia coli, and zymogen activation, zymogen degradation, and trypsin autolysis were studied. Enterokinase activated Asn-21-Tg approximately 2-fold better than Ile-21-Tg or Thr-21-Tg, and catalytic parameters of trypsins were comparable. At 37 degrees C, in 5 mm Ca(2+), all three trypsins were highly stable. In the absence of Ca(2+), Asn-21- and Ile-21-trypsins suffered autolysis in an indistinguishable manner, whereas Thr-21-trypsin exhibited significantly increased stability. In sharp contrast to observations with the rat proenzyme, at pH 8.0, 37 degrees C, autoactivation kinetics of Asn-21-Tg and Ile-21-Tg were identical; however, at pH 5. 0, Ile-21-Tg autoactivated at an enhanced rate relative to Asn-21-Tg. Remarkably, at both pH values, Thr-21-Tg showed markedly higher autoactivation rates than the two other zymogens. Finally, autocatalytic proteolysis of human zymogens was limited to cleavage at Arg-117, and no digestion at Lys-188 was detected. The observations indicate that zymogen stabilization by Ile-21 as observed in rat Tg-2 is not characteristic of human Tg-1. Instead, an increased propensity to autoactivation under acidic conditions might be relevant to the pathomechanism of the Asn-21 --> Ile mutation in hereditary pancreatitis. In the same context, faster autoactivation and increased trypsin stability caused by the Asn-21 --> Thr mutation in human Tg-1 might provide a rationale for the evolutionary divergence from Thr-21 found in other mammalian trypsinogens. (+info)Unique catalytic and molecular properties of hydrolases from Aspergillus used in Japanese bioindustries. (8/101)
This review covers the unique catalytic and molecular properties of three proteolytic enzymes and a glycosidase from Aspergillus. An aspartic proteinase from A. saitoi, aspergillopepsin I (EC 3.4.23.18), favors hydrophobic amino acids at P1 and P'1 like gastric pepsin. However, aspergillopepsin I accommodates a Lys residue at P1, which leads to activation of trypsinogens like duodenum enteropeptidase. Substitution of Asp76 to Ser or Thr and deletion of Ser78, corresponding to the mammalian aspartic proteinases, cathepsin D and pepsin, caused drastic decreases in the activities towards substrates containing a basic amino acid residue at 1. In addition, the double mutant T77D/G78(S)G79 of porcine pepsin was able to activate bovine trypsinogen to trypsin by the selective cleavage of the K6-I7 bond of trypsinogen. Deuterolysin (EC 3.4.24.39) from A. oryzae, which contains 1g atom of zinc/mol of enzyme, is a single chain of 177 amino acid residues, includes three disulfide bonds, and has a molecular mass of 19,018 Da. It was concluded that His128, His132, and Asp164 provide the Zn2+ ligands of the enzyme according to a 65Zn binding assay. Deuterolysin is a member of a family of metalloendopeptidases with a new zinc-binding motif, aspzincin, defined by the "HEXXH + D" motif and an aspartic acid as the third zinc ligand. Acid carboxypeptidase (EC 3.4.16.1) from A. saitoi is a glycoprotein that contains both N- and O-linked sugar chains. Site-directed mutagenesis of the cpdS, cDNA encoding A. saitoi carboxypeptidase, was cloned and expressed. A. saitoi carboxypeptidase indicated that Ser153, Asp357, and His436 residues were essential for the enzymic catalysis. The N-glycanase released high-mannose type oligosaccharides that were separated on HPLC. Two, which had unique structures of Man10 GlcNAc2 and Man11GlcNAc2, were characterized. An acidic 1,2-alpha-mannosidase (EC 3.2.1.113) was isolated from the culture of A. saitoi. A highly efficient overexpression system of 1,2-alpha-mannosidase fusion gene (f-msdS) in A. oryzae was made. A yeast mutant capable of producing Man5GlcNAc2 human-compatible sugar chains on glycoproteins was constructed. An expression vector for 1,2-alpha-mannosidase with the "HDEL" endoplasmic reticulum retention/retrieval tag was designed and expressed in Saccharomyces cerevisiae. The first report of production of human-compatible high mannose-type (Man5GlcNAc2) sugar chains in S. cerevisiae was described. (+info)Enteropeptidase, also known as enterokinase, is an enzyme that is produced by the intestinal brush border cells. Its primary function is to activate other digestive enzymes, most notably trypsinogen, which is a precursor to the digestive enzyme trypsin.
Trypsinogen is inactive until it is cleaved by enteropeptidase, which removes a small peptide from the N-terminus of the molecule, activating it and allowing it to participate in protein digestion. Enteropeptidase also plays a role in activating other zymogens, such as chymotrypsinogen and procarboxypeptidases, which are involved in the breakdown of proteins and peptides in the small intestine.
Deficiency or absence of enteropeptidase can lead to malabsorption and impaired digestion, as the activation of other digestive enzymes is hindered.
Trypsinogen is a precursor protein that is converted into the enzyme trypsin in the small intestine. It is produced by the pancreas and released into the duodenum, where it is activated by enterokinase, an enzyme produced by the intestinal mucosa. Trypsinogen plays a crucial role in digestion by helping to break down proteins into smaller peptides and individual amino acids.
In medical terms, an elevated level of trypsinogen in the blood may indicate pancreatic disease or injury, such as pancreatitis or pancreatic cancer. Therefore, measuring trypsinogen levels in the blood is sometimes used as a diagnostic tool to help identify these conditions.
2-Naphthylamine is a crystalline solid organic compound that is classified as a primary aromatic amine. Its chemical formula is C10H9N. It is an intensely orange-red to reddish-brown substance that is slightly soluble in water and more soluble in organic solvents.
2-Naphthylamine is produced by the reduction of 2-naphthol or its derivatives. Historically, it was used as an intermediate in the synthesis of azo dyes and other chemical compounds. However, due to its toxicity and carcinogenicity, its use has been largely discontinued in many industries.
Exposure to 2-Naphthylamine can occur through inhalation, skin contact, or ingestion, and it has been associated with an increased risk of bladder cancer and other health effects. Therefore, appropriate safety measures must be taken when handling this compound, including the use of personal protective equipment (PPE) such as gloves, lab coats, and eye protection.
Trypsin is a proteolytic enzyme, specifically a serine protease, that is secreted by the pancreas as an inactive precursor, trypsinogen. Trypsinogen is converted into its active form, trypsin, in the small intestine by enterokinase, which is produced by the intestinal mucosa.
Trypsin plays a crucial role in digestion by cleaving proteins into smaller peptides at specific arginine and lysine residues. This enzyme helps to break down dietary proteins into amino acids, allowing for their absorption and utilization by the body. Additionally, trypsin can activate other zymogenic pancreatic enzymes, such as chymotrypsinogen and procarboxypeptidases, thereby contributing to overall protein digestion.
Enterokinase4
- Enteropeptidase (also called enterokinase) is an enzyme produced by cells of the duodenum and is involved in digestion in humans and other animals. (wikipedia.org)
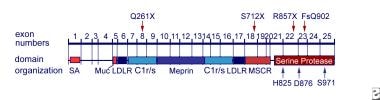
- Enteropeptidase is a serine protease (EC 3.4.21.9) consisting of a disulfide-linked heavy-chain of 82-140 kDa that anchors enterokinase in the intestinal brush border membrane and a light-chain of 35-62 kDa that contains the catalytic subunit. (wikipedia.org)
- Despite its alternative name (enterokinase), enteropeptidase is a serine protease that catalyses the hydrolysis of peptide bonds in proteins and, unlike other kinases, does not catalyze transfer of phosphate groups. (wikipedia.org)
- Enterokinase (EC 3.4.21.9) (enteropeptidase). (embl.de)
Type II transmembrane serine protease2
- Enteropeptidase is a type II transmembrane serine protease (TTSP) localized to the brush border of the duodenal and jejunal mucosa and synthesized as a zymogen, proenteropeptidase, which requires activation by duodenase or trypsin. (wikipedia.org)
- Enteropeptidase, a type II transmembrane serine protease. (medscape.com)
Serine2
- Enteropeptidase is a part of the chymotrypsin-clan of serine proteases, and is structurally similar to these proteins. (wikipedia.org)
- The activation of trypsinogen by its natural activation enzyme, the serine protease enteropeptidase, is determined by a close interaction between the two proteases. (biomedcentral.com)
Protease2
- Trypsinogen is active only after its eight-amino-acid-long activation peptide has been cleaved off by another protease, enteropeptidase. (biomedcentral.com)
- The reason is due to the advantages of adding tag on the N-terminal of the protein like enteropeptidase and TEV protease can remove N-terminal affinity tags without leaving behind any nonnative residues. (bio-bench.com)
TMPRSS151
- As the pro-region of trypsinogen contains this sequence, enteropeptidase catalyses its activation in vivo: trypsinogen → trypsin + pro-region (Val-Asp-Asp-Asp-Asp-Lys) In humans, enteropeptidase is encoded by the TMPRSS15 gene (also known as ENTK, and previously as PRSS7) on chromosome 21q21. (wikipedia.org)
Enzyme2
- This trypsinogen autoactivation process is mostly based on the fact that active trypsin can activate trypsinogen by cleaving off its activation peptide in a similar way as occurs with the activation enzyme enteropeptidase. (biomedcentral.com)
- The conversion of chymotrypsinogen to chymotrypsin is catalyzed by trypsin, which in turn arises from trypsinogen as a result of a cleavage reaction catalyzed by the enzyme enteropeptidase. (brainkart.com)
Trypsin7
- Enteropeptidase converts trypsinogen (a zymogen) into its active form trypsin, resulting in the subsequent activation of pancreatic digestive enzymes. (wikipedia.org)
- Native enteropeptidase is resistant to soybean trypsin inhibitor. (wikipedia.org)
- Enteropeptidase exhibits trypsin-like activity, cleaving proteins following a lysine at a specific cleavage site (Asp-Asp-Asp-Asp-Lys). (wikipedia.org)
- further, metformin-treated mice exhibit reduced enteropeptidase activity, reduced trypsin activity, and impaired protein digestion within the intestinal lumen. (simulations-plus.com)
- Surface-charged trypsinogen showed practically no autoactivation compared to the wild-type but could still be activated by enteropeptidase to the fully active trypsin. (biomedcentral.com)
- Trypsinogen is the inactive precursor of trypsin (PRSS1, human cationic trypsinogen), which can cleave proteins and peptides after lysine and arginine residues and is activated by the membrane-bound enteropeptidase via cleavage of the N-terminal activation peptide located on the trypsinogen surface. (biomedcentral.com)
- However, physiological conditions of course include the possibility that trypsinogen activation could also occur to a minor extent through the non-specific cleavage by proteases other than trypsin or enteropeptidase. (biomedcentral.com)
Digestive enzymes1
- Here, we report that metformin inhibits the activity of enteropeptidase and other digestive enzymes at drug concentrations predicted to occur in the human duodenum. (simulations-plus.com)
Intestinal2
- Absence of enteropeptidase results in intestinal digestion impairment. (wikipedia.org)
- Celiac disease in a patient with a congenital deficiency of intestinal enteropeptidase. (medscape.com)
Protein1
- Hoffmeister A, Dietz G, Zeitschel U, Mössner J, Rossner S, Stahl T. BACE1 is a newly discovered protein secreted by the pancreas which cleaves enteropeptidase in vitro. (medscape.com)
Deficiency2
- Some nonsense and frameshift mutations in this gene lead to a rare recessive disorder characterised by severe failure to thrive in affected infants, due to enteropeptidase deficiency. (wikipedia.org)
- Mutations in the proenteropeptidase gene are the molecular cause of congenital enteropeptidase deficiency. (medscape.com)
Trypsinogen3
- An analogous selective defect in the recognition of trypsinogen can be produced in two-chain enteropeptidase by heating or by acetylation. (wikipedia.org)
- Trypsinogen can also be autoactivated without the involvement of enteropeptidase. (biomedcentral.com)
- Besides the specific activation of trypsinogen by enteropeptidase it is well known that trypsinogen can show autoactivation [ 3 ]. (biomedcentral.com)
Specificity1
- The heavy chain influences the specificity of enteropeptidase. (wikipedia.org)
Residues1
- In the case of cattle enteropeptidase the primary translation product comprises 1035 residues with an expected mass of 114.9kDa. (wikipedia.org)
Cleavage1
- The activation cleavage site after Lys800 splits the heavy and light chains of mature cattle enteropeptidase. (wikipedia.org)
Name enteropeptidase1
- The more precise IUBMB name enteropeptidase has been in existence since 1970. (wikipedia.org)
21.91
- Vertebrate enteropeptidase ( EC 3.4.21.9 ), a type II membrane protein of the intestinal brush border, which activates trypsinogen. (embl.de)
Serine protease2
- Enteropeptidase is a type II transmembrane serine protease (TTSP) localized to the brush border of the duodenal and jejunal mucosa and synthesized as a zymogen, proenteropeptidase, which requires activation by duodenase or trypsin. (wikipedia.org)
- Enteropeptidase, a type II transmembrane serine protease. (medscape.com)
Pancreas1
- Hoffmeister A, Dietz G, Zeitschel U, Mössner J, Rossner S, Stahl T. BACE1 is a newly discovered protein secreted by the pancreas which cleaves enteropeptidase in vitro. (medscape.com)
Duodenum1
- Tripsinogen dipertahankan di pankreas lalu berpindah ke duodenum untuk dipecah menjadi tripsin aktif oleh enteropeptidase. (alomedika.com)
Proteins1
- Enteropeptidase is a part of the chymotrypsin-clan of serine proteases, and is structurally similar to these proteins. (wikipedia.org)
Active form1
- It is converted into its active form by ENTEROPEPTIDASE in the small intestine. (lookformedical.com)