The 3'-->5' exonucleases of DNA polymerases delta and epsilon and the 5'-->3' exonuclease Exo1 have major roles in postreplication mutation avoidance in Saccharomyces cerevisiae. (1/8779)
Replication fidelity is controlled by DNA polymerase proofreading and postreplication mismatch repair. We have genetically characterized the roles of the 5'-->3' Exo1 and the 3'-->5' DNA polymerase exonucleases in mismatch repair in the yeast Saccharomyces cerevisiae by using various genetic backgrounds and highly sensitive mutation detection systems that are based on long and short homonucleotide runs. Genetic interactions were examined among DNA polymerase epsilon (pol2-4) and delta (pol3-01) mutants defective in 3'-->5' proofreading exonuclease, mutants defective in the 5'-->3' exonuclease Exo1, and mismatch repair mutants (msh2, msh3, or msh6). These three exonucleases play an important role in mutation avoidance. Surprisingly, the mutation rate in an exo1 pol3-01 mutant was comparable to that in an msh2 pol3-01 mutant, suggesting that they participate directly in postreplication mismatch repair as well as in other DNA metabolic processes. (+info)Nrg1 is a transcriptional repressor for glucose repression of STA1 gene expression in Saccharomyces cerevisiae. (2/8779)
Expression of genes encoding starch-degrading enzymes is regulated by glucose repression in the yeast Saccharomyces cerevisiae. We have identified a transcriptional repressor, Nrg1, in a genetic screen designed to reveal negative factors involved in the expression of STA1, which encodes a glucoamylase. The NRG1 gene encodes a 25-kDa C2H2 zinc finger protein which specifically binds to two regions in the upstream activation sequence of the STA1 gene, as judged by gel retardation and DNase I footprinting analyses. Disruption of the NRG1 gene causes a fivefold increase in the level of the STA1 transcript in the presence of glucose. The expression of NRG1 itself is inhibited in the absence of glucose. DNA-bound LexA-Nrg1 represses transcription of a target gene 10.7-fold in a glucose-dependent manner, and this repression is abolished in both ssn6 and tup1 mutants. Two-hybrid and glutathione S-transferase pull-down experiments show an interaction of Nrg1 with Ssn6 both in vivo and in vitro. These findings indicate that Nrg1 acts as a DNA-binding repressor and mediates glucose repression of the STA1 gene expression by recruiting the Ssn6-Tup1 complex. (+info)Histone octamer transfer by a chromatin-remodeling complex. (3/8779)
RSC, an abundant, essential chromatin-remodeling complex related to SWI/SNF complex, catalyzes the transfer of a histone octamer from a nucleosome core particle to naked DNA. The newly formed octamer-DNA complex is identical with a nucleosome in all respects. The reaction requires ATP and involves an activated RSC-nucleosome intermediate. The mechanism may entail formation of a duplex displacement loop on the nucleosome, facilitating the entry of exogeneous DNA and the release of the endogenous molecule. (+info)Regulation of the start of DNA replication in Schizosaccharomyces pombe. (4/8779)
Cells of Schizosaccharomyces pombe were grown in minimal medium with different nitrogen sources under steady-state conditions, with doubling times ranging from 2.5 to 14 hours. Flow cytometry and fluorescence microscopy confirmed earlier findings that at rapid growth rates, the G1 phase was short and cell separation occurred at the end of S phase. For some nitrogen sources, the growth rate was greatly decreased, the G1 phase occupied 30-50% of the cell cycle, and cell separation occurred in early G1. In contrast, other nitrogen sources supported low growth rates without any significant increase in G1 duration. The method described allows manipulation of the length of G1 and the relative cell cycle position of S phase in wild-type cells. Cell mass was measured by flow cytometry as scattered light and as protein-associated fluorescence. The extensions of G1 were not related to cell mass at entry into S phase. Our data do not support the hypothesis that the cells must reach a certain fixed, critical mass before entry into S. We suggest that cell mass at the G1/S transition point is variable and determined by a set of molecular parameters. In the present experiments, these parameters were influenced by the different nitrogen sources in a way that was independent of the actual growth rate. (+info)The prokaryotic beta-recombinase catalyzes site-specific recombination in mammalian cells. (5/8779)
The development of new strategies for the in vivo modification of eukaryotic genomes has become an important objective of current research. Site-specific recombination has proven useful, as it allows controlled manipulation of murine, plant, and yeast genomes. Here we provide the first evidence that the prokaryotic site-specific recombinase (beta-recombinase), which catalyzes only intramolecular recombination, is active in eukaryotic environments. beta-Recombinase, encoded by the beta gene of the Gram-positive broad host range plasmid pSM19035, has been functionally expressed in eukaryotic cell lines, demonstrating high avidity for the nuclear compartment and forming a clear speckled pattern when assayed by indirect immunofluorescence. In simian COS-1 cells, transient beta-recombinase expression promoted deletion of a DNA fragment lying between two directly oriented specific recognition/crossing over sequences (six sites) located as an extrachromosomal DNA substrate. The same result was obtained in a recombination-dependent lacZ activation system tested in a cell line that stably expresses the beta-recombinase protein. In stable NIH/3T3 clones bearing different number of copies of the target sequences integrated at distinct chromosomal locations, transient beta-recombinase expression also promoted deletion of the intervening DNA, independently of the insertion position of the target sequences. The utility of this new recombination tool for the manipulation of eukaryotic genomes, used either alone or in combination with the other recombination systems currently in use, is discussed. (+info)The yeast dynamin-like protein, Mgm1p, functions on the mitochondrial outer membrane to mediate mitochondrial inheritance. (6/8779)
The mdm17 mutation causes temperature-dependent defects in mitochondrial inheritance, mitochondrial morphology, and the maintenance of mitochondrial DNA in the yeast Saccharomyces cerevisiae. Defects in mitochondrial transmission to daughter buds and changes in mitochondrial morphology were apparent within 30 min after shifting cells to 37 degrees C, while loss of the mitochondrial genome occurred after 4-24 h at the elevated temperature. The mdm17 lesion mapped to MGM1, a gene encoding a dynamin-like GTPase previously implicated in mitochondrial genome maintenance, and the cloned MGM1 gene complements all of the mdm17 mutant phenotypes. Cells with an mgm1-null mutation displayed aberrant mitochondrial inheritance and morphology. A version of mgm1 mutated in a conserved residue in the putative GTP-binding site was unable to complement any of the mutant defects. It also caused aberrant mitochondrial distribution and morphology when expressed at high levels in cells that also contained a wild-type copy of the gene. Mgm1p was localized to the mitochondrial outer membrane and fractionated as a component of a high molecular weight complex. These results indicate that Mgm1p is a mitochondrial inheritance and morphology component that functions on the mitochondrial surface. (+info)All 16 centromere DNAs from Saccharomyces cerevisiae show DNA curvature. (7/8779)
All 16 centromere DNA regions of Saccharomyces cerevisiae including 90 bp framing sequences on either side were cloned. These 300 bp long centromere regions were analysed by native polyacrylamide gel electrophoresis and found to display a reduced mobility indicative of DNA curvature. The degree of curvature is centromere dependent. The experimental data were confirmed by computer analysis of the 3-dimensional structure of the CEN DNAs. Altogether these data provide further evidence for a model for budding yeast centromeres in which CEN DNA structure could be important for the assembly, activity and/or regulation of the centromere protein-DNA complex. (+info)The nuclear ribosomal DNA intergenic spacer as a target sequence to study intraspecific diversity of the ectomycorrhizal basidiomycete Hebeloma cylindrosporum directly on pinus root systems. (8/8779)
Polymorphism of the nuclear ribosomal DNA intergenic spacer (IGS) of the ectomycorrhizal basidiomycete Hebeloma cylindrosporum was studied to evaluate whether this sequence could be used in field studies to estimate the diversity of strains forming mycorrhizas on individual Pinus pinaster root systems. This sequence was amplified by PCR from 125 haploid homokaryotic strains collected in 14 P. pinaster stands along the Atlantic coast of France by using conserved oligonucleotide primers. Restriction enzyme digestion of the amplified 3.4-kbp-long IGS allowed us to characterize 24 alleles whose frequencies differed. Nine of these alleles were found only once, whereas about 60% of the strains contained four of the alleles. Local populations could be almost as diverse as the entire population along a 150-km stretch of coastline that was examined; for example, 13 alleles were found in a single forest stand. The IGS from one strain was partially sequenced, and the sequence data were used to design oligonucleotides which allowed separate PCR amplification of three different segments of the IGS. Most polymorphisms observed among the full-length IGS regions resulted from polymorphisms in an internal ca. 1,500-bp-long sequence characterized by length variations that may have resulted from variable numbers of a T2AG3 motif. This internal polymorphic sequence could not be amplified from the genomes of nine other Hebeloma species. Analysis of this internal sequence amplified from the haploid progenies of 10 fruiting bodies collected in a 70-m2 area resulted in identification of six allelic forms and seven distinct diplotypes out of the 21 possible different combinations. Moreover, optimization of the PCR conditions resulted in amplification of this sequence from more than 80% of the DNA samples extracted from individual H. cylindrosporum infected P. pinaster mycorrhizal root tips, thus demonstrating the usefulness of this sequence for studying the below-ground diversity of mycorrhizas formed by genets belonging to the same fungal species. (+info) Fungal DNA barcoding
Fungal DNA barcoding
DNA barcoding Microbial DNA barcoding Pollen DNA barcoding DNA barcoding in diet assessment Consortium for the Barcode of Life ... as primary fungal DNA barcode, the necessity of establishing a second DNA barcode marker was expressed. Several attempts were ... Fungal DNA barcoding can help to identify and associate anamorphic and teleomorphic stages of fungi, and through that to reduce ... Fungal DNA barcoding is the process of identifying species of the biological kingdom Fungi through the amplification and ...
Fungal Diversity Survey
... and to collect and document fungi for DNA barcoding. Fungal Diversity Survey, Inc. is a Charitable 501(c)(3) organization ... Fungi Magazine, 101, pp.28-36 FunDiS , Fungal Diversity Survey Fungal Diversity Database on iNaturalist (Accuracy disputes from ... Fungal Diversity Database - Build a database of fungal observations on iNaturalist of sufficient scale and quality to be useful ... Fungal Diversity Survey, or FunDiS, is a nonprofit citizen science organization formerly known as North American Mycoflora ...
Geotrichum candidum
Alper, I; Michel, F; Labrie, S (2011). "Ribosomal DNA Polymorphisms in the Yeast Geotrichum candidum". Fungal Biology. 115 (12 ... Fungal plant pathogens and diseases, Fungal citrus diseases, Animal fungal diseases, Organisms breaking down plastic, Fungi ... Fungal growth can be supported by D-glucose, D-mannose, D-xylose, L-sorbose, D-fructose, D-galactose, sucrose, D-mannitol, D- ...
Callistosporiaceae
All species form agaricoid basidiocarps (gilled mushrooms). The family is based on recent DNA research. Vizzini A, Consiglio G ... nov., and Pleurocollybia incorporated into Callistosporium". Fungal Diversity. 101: 211-259. doi:10.1007/s13225-020-00441-x. v ...
Allomyces
... based on nuclear ribosomal DNA". Fungal Biology. 115: 381-392. doi:10.1016/j.funbio.2011.02.004. 6. Allomyces. Phillip W. ...
Physodermataceae
... based on nuclear ribosomal DNA". Fungal Biology. 115 (4): 381-392. doi:10.1016/j.funbio.2011.02.004. ISSN 1878-6146. PMID ...
Reference genome
As a result of how the DNA samples were processed, about 80 percent of the reference genome came from eight people and one male ... "1000 Fungal Genomes Project". mycocosm.jgi.doe.gov. Retrieved 2022-08-18. Genome Reference Consortium (Genome projects, ... As they are assembled from the sequencing of DNA from a number of individual donors, reference genomes do not accurately ... Instead a reference provides a haploid mosaic of different DNA sequences from each donor. For example, one the most recent ...
Point mutation
DNA replication occurs when one double-stranded DNA molecule creates two single strands of DNA, each of which is a template for ... Because RIP is so efficient at detecting and mutating repeats, fungal biologists often use it as a tool for mutagenesis. A ... These reactants can lead to both single-stranded DNA breaks and double-stranded DNA breaks. Third, bonds in DNA eventually ... It is a DNA methyltransferease-like protein, that when mutated or knocked out results in loss of RIP. Deletion of the rid ...
Blastocladiomycota
Molecular phylogeny of the Blastocladiomycota (Fungi) based on nuclear ribosomal DNA. Fungal Biology 115:381-392. Kendrick, ... Fungal Diversity. 90 (1): 135-159. doi:10.1007/s13225-018-0401-0. Hibbett DS et al. 2007. A higher-level phylogenetic ...
Physoderma
Molecular phylogeny of the Blastocladiomycota (Fungi) based on nuclear ribosomal DNA. Fungal Biology 115: 381-392. James, T.Y ...
Phylloporus
... morphological and multilocus DNA sequence analyses". Fungal Diversity. 58 (1): 73-101. doi:10.1007/s13225-012-0184-7. Neves MA ... Fungal Diversity. 55 (1): 109-123. doi:10.1007/s13225-012-0154-0. "Phylloporus on Boletineae: Surveys and Revisions". " ...
Phylloporus leucomycelinus
... morphological and multilocus DNA sequence analyses". Fungal Diversity. 58: 73-101. doi:10.1007/s13225-012-0184-7. S2CID ... Fungal Diversity. 55 (1): 109-23. doi:10.1007/s13225-012-0154-0. S2CID 17272328. Zheng NK, Tang LP, Li YC, Tolgor B, Zhu XT, ...
Fungal isolate
2005). "Structural analysis of epolactaene derivatives as DNA polymerase inhibitors and anti-inflammatory compounds". Int J Mol ... The fungal isolate ergothioneine is actively absorbed and concentrated by the human body via SLC22A4. Other notable fungal ... The statins lovastatin, mevastatin, and simvastatin precursor monacolin J, are fungal isolates. Additional fungal isolates that ... are antibiotic fungal isolates. Antibiotics retapamulin, tiamulin, and valnemulin are derivatives of the fungal isolate ...
Skeletocutis
This species is part of the community of fungal successors of decaying wood. A Finnish study found that it fruited most ... "Phylogenetic relationships in European Ceriporiopsis species inferred from nuclear and mitochondrial ribosomal DNA sequences". ... Yu, Chang-Jun; Zuo, Li; Dai, Yu-Cheng (2005). "Three polypores from Xizang new to China" (PDF). Fungal Science. 20 (3-4): 61-68 ... "Skeletocutis nothofagi Rajchenb". The Global Fungal Red List Initiative. IUCN. Retrieved 30 September 2016. Fries, Elias Magnus ...
Agaricus
"Phylogeny of the genus Agaricus inferred from restriction analysis of enzymatically amplified ribosomal DNA". Fungal Genet Biol ... Based on DNA analysis of ITS1, ITS2, and 5.8S sequences, the studied species of this section could be divided into six distinct ... The fungal genus Agaricus as late as 2008 was believed to contain about 200 species worldwide but since then, molecular ...
Dimargaritales
in Fungal Diversity vol.90, Issue 1 on page 151 in 2018. It is parasitic. But can grow solitary on agar media, thus cold ... based on nuclear small subunit ribosomal DNA sequences". Mol. Phylogenet. Evol. 16 (2): 253-62. doi:10.1006/mpev.2000.0775. ... Fungal Diversity. 90 (1): 135-159. doi:10.1007/s13225-018-0401-0. Tanabe Y, O'Donnell K, Saikawa M, Sugiyama J (August 2000). " ...
Galzinia
Li GJ, Hyde KD, Zhao RL, Hongsanan S, Abdel-Aziz FA (2016). "Fungal diversity notes 253-366" (PDF). Fungal Divers. 78. doi: ... Molecular research, based on cladistic analysis of DNA sequences, suggests the genus may be polyphyletic. The type species has ...
Nigrospora sphaerica
Fungal spores found in the body of the nail resembled the characteristic morphology of N. sphaerica. DNA sequence analysis ... Fungal spores trapped on the agar strips were developed and counted. They were then cultured into isolates allowing for ... A woman in south India was diagnosed with a fungal corneal ulcer after being hit in the eye from a cow's tail. Analysis of ... It was hypothesized that this special case of fungal corneal ulcer was caused by transfer of spores to the patients eye from ...
Lactarius
Ochricompactae" (PDF). Fungal Diversity. 28: 15-40. Eberhardt U, Verbeken A (2004). "Sequestrate Lactarius species from ... Russulales; Basidiomycota) in Alaska, based on soil and sporocarp DNA" (PDF). Molecular Ecology. 18 (10): 2213-2227. doi: ... Rinaldi AC, Comandini O, Kuyper TW (2008). "Ectomycorrhizal fungal diversity: separating the wheat from the chaff" (PDF). ... Twieg BD, Durall DM, Simard SW (2007). "Ectomycorrhizal fungal succession in mixed temperate forests". New Phytologist. 176 (2 ...
Fusarium subglutinans
DNA sequencing data reveals the presence of two major groups representing cryptic species in F. subglutinans. These were ... Fusarium subglutinans is a fungal plant pathogen. Fusarium subglutinans is the anamorph of Gibberella fujikuroi. Fusarium ... Fungal plant pathogens and diseases, Maize diseases, Mango tree diseases, Fungi described in 1925, All stub articles, ...
Savoryellaceae
"Family placement of Ascotaiwania and Ascolacicola based on DNA sequences from the large subunit rRNA gene". Fungal Divers. 2: ... Fungal Divers. 35: 1-187. E. B. Gareth Jones and Ka-Lai Pang (Editors) Marine Fungi: and Fungal-like Organisms (2012), p. 37, ... Lastly, anamorphic fungal genus Monotosporella was also added later to the order and family. Generally, Savoryellaceae species ... doi:10.1016/S0007-1536(69)80169-5. Tsui, C. K. M.; Hyde, K. D. (2003). "Freshwater mycology". Fungal Divers. Res. Ser. 10: 1- ...
Fungal contamination of contact lenses
... or DNA-based tests." Pathogenesis of the fungal contaminants include a wide range of factors such as "invasiveness, ... The issues of fungal identification observed as mentioned in this study do not pertain only to fungal contamination of contact ... "fungal strain isolated in culture." In one instance, a fungal strain previously identified as Arthrobotrys oligospora in a ... is amphotericin B which is a specific anti-fungal drug. For the treatment for filamentous fungal keratitis, "topical natamycin ...
Myriococcum
DNA analysis has shown this belongs within the Agaricales. Several other species formerly referred to Myriococcum are now ... Koukol O (2016). "Myriococcum revisited: a revision of an overlooked fungal genus". Plant Systematics and Evolution. 302 (8): ...
Copper fist
... is an N-terminal domain involved in copper-dependent DNA binding. It is named for its resemblance to a fist. It can ... be found in some fungal transcription factors. These proteins activate the transcription of the metallothionein gene in ... which allows DNA binding. Copper fist definition Turner RB, Smith DL, Zawrotny ME, Summers MF, Posewitz MC, Winge DR (July 1998 ...
Doassansiopsis
Vánky, K. (2001). "The emended Ustilaginaceae of the modern classificatory system for smut fimgi". Fungal Diversity. 6: 131-147 ... "Phylogenetic studies on nuclear large subunit ribosomal DNA sequences of smut fungi and related taxa". Canadian Journal of ...
Dolichousnea
Lücking, Robert; Nadel, Miko; Araujo, Elena; Gerlach, Alice (2020). "Two decades of DNA barcoding in the genus Usnea ( ... Fungal Diversity. 84 (1): 101-117. doi:10.1007/s13225-017-0379-z. S2CID 256071368. Ohmura 2001, p. 8. Ohmura 2001, p. 9. Ohmura ... Parmeliaceae): how useful and reliable is the ITS?". Plant and Fungal Systematics. 65 (2): 303-357. doi:10.35535/pfsyst-2020- ...
Polyploidy
Although the replication and transcription of DNA is highly standardized in eukaryotes, the same cannot be said for their ... Besides plants and animals, the evolutionary history of various fungal species is dotted by past and recent whole-genome ... Lidzbarsky GA, Shkolnik T, Nevo E (June 2009). Idnurm A (ed.). "Adaptive response to DNA-damaging agents in natural ... April 2009). "Genome sequence of the lager brewing yeast, an interspecies hybrid". DNA Research. 16 (2): 115-129. doi:10.1093/ ...
Phytophthora alni
Fungal Genet. Biol. 43 (7): 511-29. doi:10.1016/j.fgb.2006.02.006. PMID 16626980. S2CID 43275127. Chandelier A, Abras S, ... "Genetic characterization of the natural hybrid species Phytophthora alni as inferred from nuclear and mitochondrial DNA ...
Rebecca Yahr
2022) What do the first 597 global fungal red list assessments tell us about the threat status of fungi? Diversity 14 (9), 736 ... 2012) Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proceedings of ... V., Spouge, J.L., Levesque, C.A., … and Fungal Barcoding Consortium. ( ...
Pleurotheciaceae
"Fungal diversity notes 603-708: taxonomic and phylogenetic notes on genera and species". Fungal Diversity. 87 (1): 1-235. doi: ... 2015) introduced P. garethjonesii based on DNA sequence data and morphology and placed this species in Melanommataceae in ... Fungal Diversity. 87 (2): 237-298. doi:10.1007/s13225-017-0389-x. S2CID 256070993. Tsui, C. K. M.; Berbee, M. L. (2006). " ... a fungal infection of the cornea), as well as Pleurothecium recurvatum (formerly Carpoligna pleurothecii). A multi-locus ...
Fungal DNA barcoding - Wikipedia
DNA barcoding Microbial DNA barcoding Pollen DNA barcoding DNA barcoding in diet assessment Consortium for the Barcode of Life ... as primary fungal DNA barcode, the necessity of establishing a second DNA barcode marker was expressed. Several attempts were ... Fungal DNA barcoding can help to identify and associate anamorphic and teleomorphic stages of fungi, and through that to reduce ... Fungal DNA barcoding is the process of identifying species of the biological kingdom Fungi through the amplification and ...
 Target Genes, Primer Sets, and Thermocycler Settings for Fungal DNA Amplification | Fungal Diseases | CDC
Target Genes, Primer Sets, and Thermocycler Settings for Fungal DNA Amplification | Fungal Diseases | CDC
This tool assumes that fungal DNA already exists; it does not describe the procedure for purification of fungal DNA. ... Target Genes, Primer Sets, and Thermocycler Settings for Fungal DNA Amplification. *Anamorph and Teleomorph Names for Candida ... Target Genes, Primer Sets, and Thermocycler Settings for Fungal DNA Amplification. ... Trichophyton species DNA is amplified very poorly by the ITS primer set used for most other molds. There is a special set of ...
Fungal DNA Challenge in Human STR Typing of Bone Samples
The present study focuses on possible cross-reaction of fungal DNA with human STR primers that may affect subsequent forensic ... DNA analysis of forensic samples. Specificity of human STR markers namely HUMAMEL, HUMCSF1PO, D8S306, HUMTH01, HUMvWA, HUMFES/ ... amplification was inhibited in the presence of a high proportion of fungal DNA compared to human DNA (1000 ng: 1 ng) in DNA ... The present study focuses on possible cross-reaction of fungal DNA with human STR primers that may affect subsequent forensic ...
 AllPrep Bacterial/Fungal DNA/RNA/Protein Kits
AllPrep Bacterial/Fungal DNA/RNA/Protein Kits
For extraction of nucleic acids from bacterial and fungal cell cultures ... DNA & RNA Purification. Multianalyte & Virus. AllPrep Bacterial/Fungal.... AllPrep Bacterial/Fungal DNA/RNA/Protein Kits. For ... Comparison of Allprep Bacterial/Fungal DNA/RNA/Protein Kits. Features. Allprep Bacterial DNA/RNA/Protein Kit. Allprep Fungal ... Easy-to-use AllPrep Bacterial/Fungal DNA/RNA/Protein Kits isolate total nucleic acids and cellular proteins from Gram (+/-) ...
 Fungal DNA, Cells Found in Human Tumors - 3 Quarks Daily
Fungal DNA, Cells Found in Human Tumors - 3 Quarks Daily
... reports detecting fungal DNA or cells-typically at low abundance-in 35 different cancer types, with fungal species composition ... Fungal DNA, Cells Found in Human Tumors. Posted on Friday, Oct 7, 2022 7:29AM. by Azra Raza ... Fungal DNA is present in various types of cancer, according to two studies published yesterday (September 29) in Cell. The ...
 Natural contaminants in bee pollen: DNA metabarcoding as a tool to identify floral sources of pyrrolizidine alkaloids and...
Natural contaminants in bee pollen: DNA metabarcoding as a tool to identify floral sources of pyrrolizidine alkaloids and...
Keywords: Bee pollen; DNA metabarcoding; Floral diversity; Fungal diversity; LC-MS/MS; Pyrrolizidine alkaloids. ... DNA metabarcoding as a tool to identify floral sources of pyrrolizidine alkaloids and fungal diversity Food Res Int. 2021 Aug: ... Thus, a DNA metabarcoding approach based on internal transcribed spacer 2 (ITS2) region was used to identify the plant sources ... suggest that the DNA metabarcoding could be applied in a multidisciplinary approach to give a picture of floral and fungal ...
Sre1, an iron-modulated GATA DNA-binding protein of iron-uptake genes in the fungal pathogen Histoplasma capsulatum
... ... Sre1p specifically bound DNA containing the 5-(G/A)ATC(T/A)GATAA-3 sequence, and that binding was both iron- and zinc- ... The loss of iron led to a approximately 2.5-fold decrease in DNA-binding affinity, indicating that iron was directly involved ... About 0.5-1 equiv of Fe (3+) per monomer was necessary for full DNA-binding activity. Mutations in the conserved cysteine ...
 Fungi in the Antarctic cryosphere: Using DNA metabarcoding to reveal fungal diversity in glacial ice from the Antarctic...
Fungi in the Antarctic cryosphere: Using DNA metabarcoding to reveal fungal diversity in glacial ice from the Antarctic...
We assessed fungal diversity present in glacial from the Antarctic Peninsula using DNA metabarcoding through high-throughput ... 2022 Fungi in the Antarctic cryosphere: Using DNA metabarcoding to reveal fungal diversity in glacial ice from the Antarctic ... Fungi in the Antarctic cryosphere: Using DNA metabarcoding to reveal fungal diversity in glacial ice from the Antarctic ... We detected a total of 353,879 fungal DNA reads, representing 94 genera and 184 taxa, in glacial ice fragments obtained from ...
 DNA metabarcoding uncovers fungal diversity in soils of protected and non-protected areas on Deception Island, Antarctica -...
DNA metabarcoding uncovers fungal diversity in soils of protected and non-protected areas on Deception Island, Antarctica -...
DNA metabarcoding uncovers fungal diversity in soils of protected and non-protected areas on Deception Island, Antarctica. DNA ... We assessed soil fungal diversity at two sites on Deception Island, South Shetland Islands, Antarctica using DNA metabarcoding ... DNA metabarcoding of fungal diversity in air and snow of Livingston Island, South Shetland Islands, Antarctica.. ... We detected 346 fungal amplicon sequence variants dominated by the phyla Ascomycota, Basidiomycota, Mortierellomycota and ...
 Fungal DNA, allergens, mycotoxins and associations with asthmatic symptoms among pupils in schools from Johor Bahru, Malaysia. ...
Fungal DNA, allergens, mycotoxins and associations with asthmatic symptoms among pupils in schools from Johor Bahru, Malaysia. ...
DNA, Fungal | Profiles RNS
Fungal" by people in this website by year, and whether "DNA, Fungal" was a major or minor topic of these publications. ... "DNA, Fungal" is a descriptor in the National Library of Medicines controlled vocabulary thesaurus, MeSH (Medical Subject ... Below are the most recent publications written about "DNA, Fungal" by people in Profiles. ... Below are MeSH descriptors whose meaning is more general than "DNA, Fungal". ...
 Fungal Infections in Preterm Infants: Introduction, Pathogenesis, Risk Factors
Fungal Infections in Preterm Infants: Introduction, Pathogenesis, Risk Factors
Introduction The risk for invasive fungal infections is high in very low birth weight (VLBW) infants (< 1500 g) and highest for ... Examples include PCR and DNA microarray technology. The hope is that these techniques will allow for the rapid detection of ... Culture (aerobic or fungal). Culture (aerobic or fungal) or biopsy. Fungal culture or biopsy. ... Culture (aerobic or fungal). Culture (aerobic or fungal) or biopsy. Fungal culture or biopsy. ...
 Frontiers | Appendectomy Is Associated With Alteration of Human Gut Bacterial and Fungal Communities
Frontiers | Appendectomy Is Associated With Alteration of Human Gut Bacterial and Fungal Communities
... the abundance correlation networks in samples from HwA displayed more complex fungal-fungal and fungal-bacterial community ... the abundance correlation networks in samples from HwA displayed more complex fungal-fungal and fungal-bacterial community ... We assessed the alterations in gut bacterial and fungal populations associated with a history of appendectomy. In this cross- ... We assessed the alterations in gut bacterial and fungal populations associated with a history of appendectomy. In this cross- ...
 Microorganisms | Free Full-Text | Fungal Diversity Analysis of Grape Musts from Central Valley-Chile and Characterization of...
Microorganisms | Free Full-Text | Fungal Diversity Analysis of Grape Musts from Central Valley-Chile and Characterization of...
However, little is known about the diversity of fungal communities associated with grape musts, even though they are the source ... Unsurprisingly, Saccharomyces represented the most abundant fungal genus in unfermented and fermented musts, mainly due to the ... Unfermented musts were highly variable between seasons and showed higher values of fungal diversity than fermented musts. Since ... ITS PCR amplifications were carried out in 25 μL volumes containing 200 ng (~ 2 μL) of fungal DNA, 12.5 μL of MixGoTaq Green ( ...
Delivering on Antimicrobial Resistance Agenda Not Possible without Improving Fungal Diagnostic Capabilities - Volume 23, Number...
The use of new, highly sensitive, DNA detection assays (e.g., Xpert MTB/RIF) directly on respiratory specimens has transformed ... Diagnosis of fungal asthma relies on total and fungal-specific IgE testing or skin-prick testing, which is simple to perform ... Accurate Diagnosis of Fungal Sepsis in Hospitals Misdiagnosis of Smear-Negative Pulmonary Tuberculosis as Tuberculosis Fungal ... The term fungal asthma has recently been introduced as a catch-all for asthma exacerbated by fungal sensitization, airways ...
 Impact of a resistance gene against a fungal pathogen on the plant host residue microbiome: the case of the Leptosphaeria...
Impact of a resistance gene against a fungal pathogen on the plant host residue microbiome: the case of the Leptosphaeria...
DNA extraction, PCR and Illumina sequencing. Total DNA was extracted with the DNeasy Plant Mini kit (Qiagen, France), according ... Metabarcoding analysis identified 610 bacterial ASVs and 335 fungal ASVs. Fungal richness (Shannon index) was higher in Darmor- ... This fungal species has received much less attention than L. maculans, to which it is related. These two pathogenic species ... 2016). Fungal communities in organically grown winter wheat affected by plant organ and development stage. Eur. J. Plant Pathol ...
 A rust fungal effector binds plant DNA and modulates transcription - Cognitio
A rust fungal effector binds plant DNA and modulates transcription - Cognitio
 Fungal Ecology, Ectomycorrhizae and Biodiversity - Universität Innsbruck
Fungal Ecology, Ectomycorrhizae and Biodiversity - Universität Innsbruck
Ancient DNA reveals migrations around Fertile Crescent heartland - News - Nature Middle East
Analysis of Neolithic human DNA suggests there were two separate migrations from the Fertile Crescent heartland into Anatolia. ... Anti-fungals losing ground against fungal infections in hospitals. 15 November 2023 ... "The DNA traces of the past are finite and will one day be obliterated by time. It is our responsibility to learn as much as ... Ancient DNA from Mesopotamia suggests distinct Pre-Pottery and Pottery Neolithic migrations into Anatolia. Science 377, 6609 ( ...
 White-Nose Syndrome Surveillance | U.S. Geological Survey
White-Nose Syndrome Surveillance | U.S. Geological Survey
DNA-based detection of the fungal pathogen Geomyces destructans in soil from bat hibernacula White-nose syndrome (WNS) is an ... DNA-based detection of the fungal pathogen Geomyces destructans in soil from bat hibernacula White-nose syndrome (WNS) is an ... Fungal diseases are an emerging global problem affecting human health, food security and biodiversity. Ability of many fungal ... Fungal diseases are an emerging global problem affecting human health, food security and biodiversity. Ability of many fungal ...
 Robb Wolf - The Paleo Solution - Episode 214 - Dr. Michael Ruscio
Robb Wolf - The Paleo Solution - Episode 214 - Dr. Michael Ruscio
 Bacterial Diversity of Fungal Toenails Through PCR and Culture Techniques
Bacterial Diversity of Fungal Toenails Through PCR and Culture Techniques
DNA analysis is not just for crime scenes anymore. Molecular (DNA) diagnostics, targeting the genome of microorganisms, have ... Nail fungus treatments may be failing because what we have identified as a fungal nail is actually a mixed bacterial and fungal ... Bacterial Diversity of Fungal Toenails Through PCR and Culture Techniques. Grant Winners. *Paul Greenman, DPM - College of ... Treatment of a fungal nail is often expensive, painful, and unsuccessful. Surgery, dangerous oral medications, topical ...
 Fungal Planet description sheets: 1042-1111: Ingenta Connect
Fungal Planet description sheets: 1042-1111: Ingenta Connect
The journal publishes high-quality papers elucidating known and novel fungal taxa at the DNA level, and also strives to present ... download Fungal Planet description sheets: 1042–1111 Download (PDF 42,329.2 kb) ... This article was made available online on June 29, 2020 as a Fast Track article with title: "Fungal Planet description sheets: ... A further aim is to promote fungal taxonomy by employing a polythetic approach to clarify the true phylogeny and relationships ...
 F fungal soil communities had been much extra complicated (Fig. 1). Quite a few fungalF | DNA Alkylatingdna-alkylating.com
F fungal soil communities had been much extra complicated (Fig. 1). Quite a few fungalF | DNA Alkylatingdna-alkylating.com
1). A number of fungal ITS forms had been abundant in all replicate DNA samples from J2 of one particular or extra soils but ... 1). Quite a few fungalF fungal soil communities were substantially far more complicated (Fig. ... F fungal soil communities had been much extra complicated (Fig. ... Author: DNA_ Alkylatingdna View all posts by DNA_ Alkylatingdna ... F fungal soil communities had been much extra complicated (Fig. 1). Quite a few fungal. F fungal soil communities were ...
 Study Says Organic Milk Offers No Nutrition Boost | Inside Science
Study Says Organic Milk Offers No Nutrition Boost | Inside Science
 Synthesis, antimicrobial evaluation, DNA gyrase inhibition, and in silico pharmacokinetic studies of novel quinoline...
Synthesis, antimicrobial evaluation, DNA gyrase inhibition, and in silico pharmacokinetic studies of novel quinoline...
... and fungal strains (Aspergillus fumigatus, Syncephalastrum racemosum, Geotrichum candidum, and Candida albicans). Compounds ... Synthesis, antimicrobial evaluation, DNA gyrase inhibition, and in silico pharmacokinetic studies of novel quinoline ... Synthesis, antimicrobial evaluation, DNA gyrase inhibition, and in silico pharmacokinetic studies of novel quinoline ... Herein, we report the synthesis and in vitro antimicrobial evaluation of novel quinoline derivatives as DNA gyrase inhibitors. ...
 Determination of Genetic Divergence Based on DNA Markers Amongst Monosporidial Strains Derived from Fungal Isolates of Karnal...
Determination of Genetic Divergence Based on DNA Markers Amongst Monosporidial Strains Derived from Fungal Isolates of Karnal...
Ed [44]. Although a number of fungal species will not be only able to synthesizeEd | Dna-Pk Inhibitor dna-pkinhibitor.com
Though several fungal species will not be only capable to synthesize ABA theyre capable of sensing the hormone applying ... Although a number of fungal species will not be only able to synthesizeEd [44]. ... Ed [44]. Although a number of fungal species will not be only able to synthesize. Ed [44]. Though several fungal species will ... Already within the presence of 100 nM ABA spore germination also as fungal growth are promoted [45]. The virulence of ...
 Improving molecular detection of Candida DNA in whole blood: Comparison of seven fungal DNA extraction protocols using real...
Improving molecular detection of Candida DNA in whole blood: Comparison of seven fungal DNA extraction protocols using real...
Dive into the research topics of Improving molecular detection of Candida DNA in whole blood: Comparison of seven fungal DNA ... T1 - Improving molecular detection of Candida DNA in whole blood. T2 - Comparison of seven fungal DNA extraction protocols ... Improving molecular detection of Candida DNA in whole blood: Comparison of seven fungal DNA extraction protocols using real- ... Improving molecular detection of Candida DNA in whole blood: Comparison of seven fungal DNA extraction protocols using real- ...
 Links to the best Breeding & Whelping sites and articles on the net!
Links to the best Breeding & Whelping sites and articles on the net!
FUNGAL INFECTION. NEW DNA tests for Blastomycosis and Aspergilosis. *CANINE HERPES VIRUS INFECTION. Canine herpes virus is ... AFGHAN HOUND COAT COLORS AND DNA TESTS. A brief explanation of how currently available DNA tests can help predict some coat ... DNA SMOKESCREEN. Controversial? Yes. But undeniable common sense for top Breeders. *GENE SCIENCE YOU DONT WANT TO KNOW ABOUT. ... VETERINARY DIAGNOSTICS CENTER - DNA SERVICES FOR ANIMALS Identify carriers to enhance breed improvement programs and identify ...
FungiSpeciesBacterialDiversityInfectionsDifferences in soil fungal communitiesBarcodingGenesAmplicon sequence variantsMicrobiomeGeneraTaxaSoilSpecimensStrainsMolecularSequenceAmplificationInfectionCandidaPathogensMycotoxinsRibosomalMicrobialCompositionSoilsPrimersCommunitiesGenomicEndophytesAnalyseBarcodesSystematicsDiseasesGenomeHerbariumSystemicCysteineContaminantsDiagnosisEcologyPurificationBindsOnychomycosisSpecificityCandidiasisDistinctExtractionEpidemiologyClosely
Fungi21
- Fungal DNA barcoding is the process of identifying species of the biological kingdom Fungi through the amplification and sequencing of specific DNA sequences and their comparison with sequences deposited in a DNA barcode database such as the ISHAM reference database, or the Barcode of Life Data System (BOLD). (wikipedia.org)
- In this attempt, DNA barcoding relies on universal genes that are ideally present in all fungi with the same degree of sequence variation. (wikipedia.org)
- Fungal DNA barcoding can help to identify and associate anamorphic and teleomorphic stages of fungi, and through that to reduce the confusing multitude of fungus names. (wikipedia.org)
- The success of identification of fungi by means of DNA barcode sequences stands and falls with the quantitative (completeness) and qualitative (level of identification) aspect of the reference database. (wikipedia.org)
- 2012) proposed the ITS region as primary DNA barcode region for the genetic identification of fungi. (wikipedia.org)
- This document describes some of the target genes and primers that can be used for DNA sequence-based identification of fungi and the PCR conditions with which to use those primers. (cdc.gov)
- On the other hand, the molecular analysis allowed identifying several fungal genera in 33 samples, including the toxigenic fungi Alternaria and Aspergillus, which were positively correlated to the plant genus Hypericum. (nih.gov)
- However, the majority of the fungal diversity comprised taxa of rare and intermediate relative abundance, predominately known mesophilic fungi. (nerc.ac.uk)
- In both sampling areas the DNA of opportunistic, phytopathogenic and symbiotic fungi were detected, which might have been introduced by human activities, transported by birds or wind, and/or represent resident fungi not previously reported from Antarctica. (bas.ac.uk)
- Fungal community structure differed between the two lines at harvest, but not subsequently, suggesting that the presence/absence of the resistance gene influences the microbiome at the base of the stem whilst the plant is alive, but that this does not necessarily lead to differential colonisation of the residues by fungi. (biorxiv.org)
- The sequences corresponded to fungal ITS of eight genera of Ascomycota, five genera of Basidiomycota, Rhizopodium (Chytridiomycota), and Mortierella (Fungi incertae sedis) (Table 2). (dna-alkylating.com)
- Characterizing and understanding possible changes to, and survival of, fungal species in environments like the ILMAH is of high importance since fungi are not only potentially hazardous to the inhabitants but could also deteriorate the habitats themselves. (sciencedaily.com)
- With most fungal species yet to be discovered, the extent of the importance of fungi is most likely underestimated. (kew.org)
- Common DNA regions that are sequenced for identifying fungi to a species level include the external transcribed spacer (ETS), small ribosomal subunit (SSU), internal transcribed spacer 1 (ITS1), 5.8S ribosomal RNA sequence (5.8S), internal transcribed spacer 2 (ITS2), large ribosomal subunit (LSU), and intergenic spacer (IGS). (msu.edu)
- In this study, DNA sequence data were used to characterize 290 Fusarium strains isolated during a survey of root-colonizing endophytic fungi of agricultural and nonagricultural plants in northern Kazakhstan. (researchgate.net)
- The Venice Museum hosts the largest and best preserved fungal collection in Italy with more than 25,000 samples, representing approximately 6,000 species of fungi including many rare specimens. (berkeley.edu)
- because mushrooms are only produced seasonally and sometimes very rarely, e.g. only once every several years, the resulting database will facilitate identification of fungi present in plants, in the soil, and in the air at all times, simply by comparing DNA information with the one generated by this and other studies. (berkeley.edu)
- The DNA sequence data generated by this project are now available to the public and research communities and can enhance the ability to identify fungi in the environment. (berkeley.edu)
- DNA-based identification of fungi in the environment can aid in the diagnosis of plant and animal diseases, enhance studies of fungal diversity and evolution, and lead to a better understanding of factors influencing nutrient cycling and productivity in forests and agricultural ecosystems. (berkeley.edu)
- Overview of Fungal Infections Fungi are eukaryotic organisms that exist as yeast, molds, or both forms. (msdmanuals.com)
- The Kew databases contains data and resources concerning plants and fungi such as herbarium and fungarium catalogues, a seed information database, a DNA and tissue bank, accepted plant and fungal names, lists of synonyms and medicinal plant names, and much more. (lu.se)
Species24
- The interspecific variation, i.e., the variation between species, in the chosen DNA barcode gene should exceed the intraspecific (within-species) variation. (wikipedia.org)
- Moreover, fungal species can comprise multiple strains that can vary in their morphology or in traits such as carbon- and nitrogen utilisation, which has often led to their description as different species, eventually producing long lists of synonyms. (wikipedia.org)
- Trichophyton species DNA is amplified very poorly by the ITS primer set used for most other molds. (cdc.gov)
- One group, led by researchers at the Weizmann Institute of Science in Israel and the University of California, San Diego (UCSD), reports detecting fungal DNA or cells-typically at low abundance-in 35 different cancer types, with fungal species composition differing among them. (3quarksdaily.com)
- Fungal species boundaries in the genomics era. (uchicago.edu)
- The DNA ATM Formulation sequences of ITS kinds were determined to recognize fungal species that potentially interacted together with the J2 in soil. (dna-alkylating.com)
- Though several fungal species will not be only capable to synthesize ABA they're capable of sensing the hormone applying specific receptors. (dna-pkinhibitor.com)
- Though a number of fungal species are which acts as transcription factor and is really a crucial compound in the abscisic acid signaling not only capable to synthesize ABA they may be capable of sensing the hormone making use of distinct pathway. (dna-pkinhibitor.com)
- In order to determine which fungal species were present and how the composition of the mycobiome changed during human habitation, samples collected at various time points in a 30-day period were characterized. (sciencedaily.com)
- The researchers gene sequenced the samples to show which species of fungus were present and to determine the total (alive and dead) and viable (alive and able to reproduce) fungal populations. (sciencedaily.com)
- With the advent and advancement of DNA based molecular studies over the last few decades, species, genera, families and orders of Ascomycota have been subjected to rapid taxonomic changes. (researchgate.net)
- These fungal species were collected from more than 300 host species in terrestrial habitats of different provinces in Italy. (researchgate.net)
- Moreover, with major environmental threats such as climate change, we may be losing fungal species to extinction before we even discover them in understudied areas like Boyacá. (kew.org)
- 2017). Just a 50-day fungal inventory revealed up to 308 potentially new species not previously included in the global sequence repositories. (kew.org)
- The overall goal of this project is to answer the question of what fungal species can be found in Boyacá, implementing state-of-the-art approaches to analyse fungal diversity and developing a new protocol that is reproducible and easy to disseminate. (kew.org)
- By performing PCR and sequencing the entire ribosomal operon (~10kb) instead of sequencing each region individually through Sanger Sequencing, scientists can gain high quality data in a high throughput manner, providing clarity and resolution of fungal species relatedness. (msu.edu)
- My work as a master's student is focused on investigating the diversity of species in the fungal family Bankeraceae represented in western North America. (botanicgardens.org)
- Scientists are also starting to better understand how fungal viruses move between species. (theconversation.com)
- Traditional methods of identifying fungal exposures in occupational environments, such as culture and microscopy-based approaches, have several limitations that have resulted in the exclusion of many species. (cdc.gov)
- ITS sequencing results in the detection of many fungal species that are either not detected or difficult to identify to species level using culture or microscopy. (cdc.gov)
- While these methods do not provide quantitative measures of fungal burden, they offer a new approach to hazard identification and can be used to determine overall species richness and diversity within an occupational environment. (cdc.gov)
- DNA probes are available for several species. (medscape.com)
- Mitochondrial quality control prevents accumulation of intramitochondrial-derived reactive oxygen species (mtROS), thereby protecting cells against DNA damage, genome instability, and programmed cell death. (lu.se)
- However, underlying mechanisms are incompletely understood, particularly in fungal species. (lu.se)
Bacterial11
- AllPrep Bacterial/Fungal. (qiagen.com)
- Easy-to-use AllPrep Bacterial/Fungal DNA/RNA/Protein Kits isolate total nucleic acids and cellular proteins from Gram (+/-) bacterial cultures and fungal cell cultures. (qiagen.com)
- Cultured bacterial and fungal cells are first lysed using glass bead tubes, and nucleic acids and proteins are solubilized and mixed with a binding solution. (qiagen.com)
- We assessed the alterations in gut bacterial and fungal populations associated with a history of appendectomy. (frontiersin.org)
- Compared with those in samples from HwoA, the abundance correlation networks in samples from HwA displayed more complex fungal-fungal and fungal-bacterial community interactions. (frontiersin.org)
- Direct interactions with other members of the community involved many fungal and bacterial ASVs (amplicon sequence variants). (biorxiv.org)
- Nail fungus treatments may be failing because what we have identified as a fungal nail is actually a mixed bacterial and fungal infection (polymicrobial onychopathy). (nova.edu)
- A subsequent in vitro investigation against the bacterial DNA gyrase target enzyme revealed a significant potent inhibitory activity of quinoline derivative 14, which can be observed from its IC50 value (3.39 μM). (olmdiagnostics.com)
- Mycetoma has two distinct etiologies: fungal and bacterial. (who.int)
- The resulting immune system abnormalities make people with AD-HIES highly susceptible to infections, particularly bacterial and fungal infections of the lungs and skin. (medlineplus.gov)
- Uncontrolled and/or active systemic infection (viral, bacterial or fungal) b. (who.int)
Diversity13
- Thus, a DNA metabarcoding approach based on internal transcribed spacer 2 (ITS2) region was used to identify the plant sources of 17 PAs/PANOs detected by a validated method in liquid chromatography coupled to mass spectrometry (LC-MS/MS), as well as floral and fungal diversity in 61 bee pollen samples. (nih.gov)
- Despite limitations in identifying some pollen types, these preliminary results suggest that the DNA metabarcoding could be applied in a multidisciplinary approach to give a picture of floral and fungal diversity, which can be sources of natural contaminants in bee pollen and would help to control its safety. (nih.gov)
- We assessed fungal diversity present in glacial from the Antarctic Peninsula using DNA metabarcoding through high-throughput sequencing (HTS). (nerc.ac.uk)
- We assessed soil fungal diversity at two sites on Deception Island, South Shetland Islands, Antarctica using DNA metabarcoding analysis. (bas.ac.uk)
- DNA metabarcoding of fungal diversity in air and snow of Livingston Island, South Shetland Islands, Antarctica. (bas.ac.uk)
- We showed that the overall fungal diversity changed when humans were present. (sciencedaily.com)
- They showed that the diversity of the mycobiome and the levels of different fungal populations changed during the duration of the experiment. (sciencedaily.com)
- However, to be able to show that increased fungal diversity is a result of human presence, the mycobiome of the occupants will also need to be studied. (sciencedaily.com)
- The goal of the project described here is to assess the unknown fungal diversity in the department (region) of Boyacá in Colombia, and will run for 10 months until March 2018. (kew.org)
- Fungal research in Colombia could unveil similar diversity patterns as a recent study in southern South America (Truong et al. (kew.org)
- Moreover, at a time when fungal biologists and mycology training are decreasing in most academic institutions, we seek to provide training of local researchers in new techniques as well as in general fungal diversity and taxonomy. (kew.org)
- Overall, straw retention improved α-diversity, network structure and fungal community, while reduced soil pathogenic microbes across the entire soil profile. (biomedcentral.com)
- This protocol details an approach for determining fungal diversity within air samples through genomic DNA extraction, amplification, sequencing, and taxonomic identification of fungal internal transcribed spacer (ITS) regions. (cdc.gov)
Infections14
- Prior to studies of and broad institution of antifungal prophylaxis in high-risk preterm infants, the incidence of fungal infections had been rising in infants born at less than 1000 g, with the ensuing resuscitation and survival of more and more infants. (medscape.com)
- For these reasons, fungal infections are often difficult to eradicate in the preterm infant and, in cases of candidemia, central venous catheter removal is critical for clearance and survival. (medscape.com)
- Although these immunocompromised infants are at increased risk during most of their hospital stay, they are at the highest risk of acquiring invasive fungal infections during the first weeks of life, when the most invasive therapies are performed and remain in place. (medscape.com)
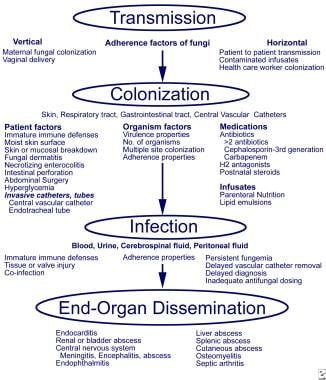
- The pathogenesis of fungal infections in preterm infants involves adherence, colonization, and dissemination (as shown in the image below). (medscape.com)
- Pathogenesis and invasive fungal infections in very low birth weight infants. (medscape.com)
- Furthermore, the importance of the accurate and timely diagnosis of fungal infections in defeating AMR has been starkly absent from policy discussions ( 4 ). (cdc.gov)
- We provide 4 examples of specific clinical situations that require greater application of existing fungal diagnostics and improved overall fungal diagnostic capability and are in line with the 95-95 by 2025 Roadmap from the Global Action Fund for Fungal Infections ( 6 ). (cdc.gov)
- Also, while secondary external fungal infections are relatively common in teleosts and elasmobranchs, primary systemic fungal infections are rarely reported. (vin.com)
- For these reasons, fungal infections are often difficult to eradicate in the preterm infant. (medscape.com)
- This article, focuses on noncandidal oral fungal infections (deep mycoses). (medscape.com)
- Although these noncandidal fungal infections are considerably less common than oral candidiasis, they commonly produce subclinical infection, especially pulmonary infections. (medscape.com)
- The oral lesions associated with these deep fungal infections are chronic and progress to form solitary, chronic deep ulcers with the potential for local destruction and invasion and systemic dissemination. (medscape.com)
- Illness in a person with compelling evidence (e.g., culture, histopathology, seroconversion) of a different fungal infection, such as histoplasmosis or coccidioidomycosis, and meeting only non-confirmatory laboratory criteria for blastomycosis should not be counted as a case of blastomycosis since other fungal infections can cause false positive Blastomyces antigen and antibody test results. (cdc.gov)
- The fungal superbug Candida auris causes serious and often fatal infections. (cdc.gov)
Differences in soil fungal communities2
- Furthermore, no-tillage and traditional agricultural systems have significant differences in soil fungal communities [ 25 ], but little is known about fungal communities' response to soil depth in sugarcane cultivation system. (biomedcentral.com)
- As black spruce stands are also covered by ericaceous shrubs, we investigated if differences in soil fungal communities and ericaceous shrubs abundance could explain the differences observed in balsam fir growth and nutrition. (frontiersin.org)
Barcoding2
- Another crucial prerequisite for DNA barcoding is the ability to unambiguously trace the provenance of DNA barcode data back to the originally sampled specimen, the so-called voucher specimen. (wikipedia.org)
- When the identity of a certain taxon (or a genetic sequence in the case of DNA barcoding) is in doubt, the original specimen can be re-examined to review and ideally solve the issue. (wikipedia.org)
Genes3
- The loss of iron led to a approximately 2.5-fold decrease in DNA-binding affinity, indicating that iron was directly involved in SRE1 regulation of iron-uptake genes. (nih.gov)
- PIF1 family DNA helicases suppress R-loop mediated genome instability at tRNA genes. (uchicago.edu)
- To carry out its roles, the STAT3 protein attaches to DNA and helps control the activity of particular genes. (medlineplus.gov)
Amplicon sequence variants1
- We detected 346 fungal amplicon sequence variants dominated by the phyla Ascomycota, Basidiomycota, Mortierellomycota and Chytridiomycota. (bas.ac.uk)
Microbiome1
- This study, from Illinois, focuses on the alteration of the Massasaugas snake skin microbiome by fungal diseases. (msu.edu)
Genera2
- We detected a total of 353,879 fungal DNA reads, representing 94 genera and 184 taxa, in glacial ice fragments obtained from seven sites in the north-west Antarctic Peninsula and South Shetland Islands. (nerc.ac.uk)
- Bands 9 and 15, of which the DNA was most closely associated to the genera Davidiella and Rhizophydium, respectively, had been connected with J2 from all 3 soils, despite the fact that they wereFIG 1 DGGE profiles of fungal ITS fragments amplified from DNA of M. hapla J2 from 3 arable soils and from total soil DNA. (dna-alkylating.com)
Taxa3
- Only 26 (14%) of the total fungal taxa detected were present at all sampling sites. (nerc.ac.uk)
- In the last eight years, we have introduced many novel fungal taxa with numerous new host and country records of ascomycetous microfungi from Italy. (researchgate.net)
- Besides, both sugarcane straw retention and soil depths (topsoil and subsoil) significantly altered the co-occurrence patterns and fungal keystone taxa closely related to straw decomposition. (biomedcentral.com)
Soil10
- F fungal soil communities had been much extra complicated (Fig. 1). (dna-alkylating.com)
- F fungal soil communities were substantially far more complicated (Fig. 1). (dna-alkylating.com)
- A number of fungal ITS forms had been abundant in all replicate DNA samples from J2 of one particular or extra soils but not in the surrounding soil, suggesting precise attachment to the J2 in soil (Fig. 1, bands two, 3, 4, 6, 9, 11, 13, and 15). (dna-alkylating.com)
- A number of the fungal ITS kinds associated with J2 have been also abundant in soil, but the relative band intensity within the profile was higher for the J2 samples than for soil, which indicated an enrichment on J2 (Fig. 1, bands 1, 5, 7, eight, ten, 12, and 14). (dna-alkylating.com)
- The most reproducible patterns had been detected on J2 from replicates with the most suppressive soil Kw, evidencing by far the most distinct fungal attachment when compared with these in the other two soils. (dna-alkylating.com)
- Collect and document fungal specimens and soil samples in the four types of habitats present in Boyacá. (kew.org)
- The purpose of this study was to provide sufficient information on soil fungal community networks and their functions in response to straw retention. (biomedcentral.com)
- Hence, we used quantitative real-time PCR (qRT-PCR), Illumina MiSeq (ITS rRNA) and FUNGuild to examine ITS rRNA gene populations, soil fungal succession and their functions under control (CK) and sugarcane straw retention (SR) treatments at different soil layers (0-10, 10-20, 20-30, and 30-40 cm) in fallow fields. (biomedcentral.com)
- Thus, retaining straw to improve fungal composition, community stability and their functions, in addition to reducing soil-borne pathogens, can be an essential agronomic practice in developing a sustainable agricultural system. (biomedcentral.com)
- High throughput sequencing (HTS) has been employed to investigate fungal community composition in soil, however, our understanding of the function and network of fungal community in a fallow straw retention system is very limited [ 24 ]. (biomedcentral.com)
Specimens4
- Voucher specimens should be clearly labelled as such, including a permanent voucher identifier that unambiguously connects the specimen with the DNA barcode data derived from it. (wikipedia.org)
- Practicing physicians will obtain nail specimens for standard laboratory and DNA analysis. (nova.edu)
- In this study, a Candida-specific real-time PCR assay was used to detect Candida albicans DNA in inoculated whole blood specimens extracted using seven different extraction protocols. (ukhsa.gov.uk)
- To do this, I am attempting to generate DNA barcode data from all 700 specimens. (botanicgardens.org)
Strains1
- The preliminary antimicrobial activity was assessed against a panel of pathogenic microbes including Gram-positive bacteria (Streptococcus pneumoniae and Bacillus subtilis), Gram-negative bacteria (Pseudomonas aeruginosa and Escherichia coli), and fungal strains (Aspergillus fumigatus, Syncephalastrum racemosum, Geotrichum candidum, and Candida albicans). (olmdiagnostics.com)
Molecular5
- Molecular (DNA) diagnostics, targeting the genome of microorganisms, have shown that there are often more and different pathogens causing nail disease than what we currently identify via cultivation in lab. (nova.edu)
- Also, a molecular docking study of the most active compounds was carried out to explore the binding affinity of the new ligands toward the active site of DNA gyrase enzyme as a proposed target of their activity. (olmdiagnostics.com)
- dPCR Microbial DNA Detection Assays are intended for molecular biology applications. (qiagen.com)
- Advances in the field over the last two decades have led occupational health researchers to turn to molecular-based approaches for identifying fungal hazards. (cdc.gov)
- CDC sequenced nearly 45,000 DNA samples by using Advanced Molecular Detection (AMD) technologies. (cdc.gov)
Sequence5
- Like the flanking 18S and 28S subunits, the 5.8S subunit contains a highly conserved DNA sequence, as they code for structural parts of the ribosome, which is a key component in intracellular protein synthesis. (wikipedia.org)
- Sre1 had sequence similarity to the fungal negative regulators of siderophore biosynthesis. (nih.gov)
- Sre1p specifically bound DNA containing the 5'-(G/A)ATC(T/A)GATAA-3' sequence, and that binding was both iron- and zinc-dependent. (nih.gov)
- In the past few decades the technology needed to sequence and study fungal viruses has improved in leaps and bounds. (theconversation.com)
- The project started in April 2006, and collection of DNA sequence data was completed in 2011. (berkeley.edu)
Amplification2
- Notably, amplification was inhibited in the presence of a high proportion of fungal DNA compared to human DNA (1000 ng: 1 ng) in DNA mixture experiments. (astm.org)
- However, the detection of low titres of Candida DNA in blood from patients with candidaemia requires the use of extraction methods that efficiently lyse yeast cells and recover small amounts of DNA suitable for amplification. (ukhsa.gov.uk)
Infection5
- All of these factors place them at high risk for fungal infection. (medscape.com)
- Within the context of stewardship programs, inadequate attention has been paid to fungal infection as the cause of antibacterial treatment failure. (cdc.gov)
- Accurate diagnosis or exclusion of fungal infection will have a substantial effect on antimicrobial drug usage and on our ability to limit AMR to bacteria. (cdc.gov)
- Candidiasis (candidosis) is by far the most common fungal infection of the mouth (oral cavity). (medscape.com)
- Cross-reactivity is a known problem with the EIA antigen test, and cases known to be infected with another fungal infection should not be counted as blastomycosis cases. (cdc.gov)
Candida1
- More importantly, the YeaStar method enabled C. albicans DNA to be detected with highest sensitivity over the entire range of copy numbers evaluated, and appears to be an optimal method for extracting Candida DNA from whole blood. (ukhsa.gov.uk)
Pathogens1
- Compounds that revealed the best activity were subjected to further biological studies to determine their minimum inhibitory concentrations (MICs) against the selected pathogens as well as their in vitro activity against the E. coli DNA gyrase, to realize whether their antimicrobial action is mediated via inhibition of this enzyme. (olmdiagnostics.com)
Mycotoxins2
- Fungal DNA, allergens, mycotoxins and associations with asthmatic symptoms among pupils in schools from Johor Bahru, Malaysia. (lu.se)
- Take fungal poisons (mycotoxins), for example. (knowthecause.com)
Ribosomal4
- The region is flanked by the DNA sequences for the ribosomal small subunit (SSU) or 18S subunit at the 5' end, and by the large subunit (LSU) or 28S subunit at the 3' end. (wikipedia.org)
- The ribosomal operon consisting of the ETS, SSU, ITS1, 5.8S, ITS2, LSU, and IGS regions typically sequenced individually as fungal barcodes, and associated primers used to amplify various regions via PCR. (msu.edu)
- These sequences are very close together, and are often found clustered in tandem repeats in fungal genomes known as a ribosomal operon (Fig. 1). (msu.edu)
- A diverse representation of the Venice collection was sampled and sent to U.C Berkeley, where a portion of the genome (the ribosomal DNA internal transcribed spacer, or ITS, region) was sequenced and analyzed. (berkeley.edu)
Microbial4
- The dPCR Microbial DNA Detection Assays and the QIAcuity dPCR System enable accurate digital PCR-based quantification. (qiagen.com)
- Our dPCR Microbial DNA Detection Assays can be combined for multiplex analysis. (qiagen.com)
- A comparison between singleplex and multiplex analysis using the dPCR Microbial DNA Detection Assays shows similar and highly precise quantification for all targets (see figure Microbial detection in multiplex dPCR on QIAcuity ). (qiagen.com)
- The dPCR Microbial DNA Detection Assays are intended for use in nanoplate digital PCR. (qiagen.com)
Composition2
- PA/PANO-producing plants were identified in 54% of the PA/PANO-contaminated samples analysed by DNA metabarcoding, which also allowed identifying the overall floral and fungal composition of 56 samples. (nih.gov)
- Human presence in closed habitats that may one day be used to explore other planets is associated with changes in the composition of the fungal community - the mycobiome - that grows on surfaces inside the habitat, according to a new study. (sciencedaily.com)
Soils1
- Fungal community DNA was sequenced from soils in northern, subarctic Sweden. (data.gov.uk)
Primers1
- The present study focuses on possible cross-reaction of fungal DNA with human STR primers that may affect subsequent forensic DNA analysis of forensic samples. (astm.org)
Communities3
- All communities should have access to nonculture fungal diagnostics, which can substantially benefit clinical outcome, antimicrobial stewardship, and control of antimicrobial resistance. (cdc.gov)
- Knowing how fungal communities change in the presence of humans is thus necessary for the development of appropriate countermeasures to maintain habitats like the ILMAH or the ISS and to protect the health of the people who live there. (sciencedaily.com)
- Our results reveal that fungal communities observed under aspen are drivers of balsam fir early growth and nutrition in boreal forest stands and may facilitate ecotone migration in a context of climate change. (frontiersin.org)
Genomic2
- The yield and quality of total nucleic acids were estimated using UV absorbance, and specific recovery of C. albicans genomic DNA was estimated quantitatively in comparison with a reference (Qiagen kit/lyticase) method currently in use in our laboratory. (ukhsa.gov.uk)
- Of the seven extraction protocols evaluated, only the MasterPure yeast DNA extraction reagent kit gave significantly higher total nucleic acid yields than the reference method, although nucleic acid purity was highest using either the reference or YeaStar genomic DNA kit methods. (ukhsa.gov.uk)
Endophytes2
- Fungal endophytes can improve plant tolerance to abiotic stress conditions. (researchgate.net)
- Fungal root endophytes, including the common form group of dark septate endophytes (DSEs), represent different taxonomic groups with potentially diverse life strategies. (researchgate.net)
Analyse1
- Lazaridis was part of a multidisciplinary study that included archaeologists, anthropologists, and bioinformatics specialists, and is the first to analyse DNA data collected from Neolithic individuals across Mesopotamia (South-eastern Turkey and Northern Iraq), Cyprus, the Zagros mountains and Armenia. (natureasia.com)
Barcodes1
- Morphological and culture characteristics are supported by DNA barcodes. (ingentaconnect.com)
Systematics1
- A fundamental problem in fungal systematics is the existence of teleomorphic and anamorphic stages in their life cycles. (wikipedia.org)
Diseases2
- Lack of routine diagnostic testing for fungal diseases exacerbates the problem of antimicrobial drug empiricism, both antibiotic and antifungal. (cdc.gov)
- Fungal viruses have been important in reducing the impact of fungal diseases on chestnuts in Europe. (theconversation.com)
Genome1
- Genome-wide DNA data collected from people who lived in the Fertile Crescent and surrounding areas during the Neolithic period is enabling scientists to piece together the likely population movements and genetic influences on early farmers in the region. (natureasia.com)
Herbarium1
- Kew databases innehåller data och resurser om växter och svampar såsom herbarium- och fungariumkataloger, en fröinformationsdatabas, en DNA- och vävnadsbank, accepterade växt- och svampnamn, listor på synonymer och medicinala växtnamn etc. (lu.se)
Systemic1
- On 25 January, surgery was performed in an attempt to biopsy the internal organs to check for systemic fungal spread, evaluate organs for predisposing conditions such as neoplasia, and remove the left uterus. (vin.com)
Cysteine1
- this response is mediated by oxidation of the cysteine residue on its DNA binding domain, which enhances DNA binding. (lu.se)
Contaminants1
- The results of the present study underscore the importance of carefully analyzing the presence of non-human biological contaminants that may affect DNA typing of environmentally challenged forensic samples to avoid spurious data interpretation. (astm.org)
Diagnosis1
- There are other useful techniques for the diagnosis of mycetoma such as DNA sequencing. (who.int)
Ecology1
- Fungal Ecology 30: 135-138. (plymouth.ac.uk)
Purification1
- it does not describe the procedure for purification of fungal DNA. (cdc.gov)
Binds1
- DNA binds to the first spin column and the flow-through is combined with a solution to bind the total RNA on a second spin column. (qiagen.com)
Onychomycosis1
- This study will test 25-50 individuals seeking treatment for suspected fungal nail disease (onychomycosis). (nova.edu)
Specificity2
- The specificity of these ten STR markers for human DNA was demonstrated. (astm.org)
- The TaqMan PCR assay used to amplify the DNA extracts achieved high levels of specificity, sensitivity and reproducibility. (ukhsa.gov.uk)
Candidiasis1
- Candidiasis is a sporadic fungal disease in livestock. (springer.com)
Distinct1
- Ancient DNA from Mesopotamia suggests distinct Pre-Pottery and Pottery Neolithic migrations into Anatolia. (natureasia.com)
Extraction1
- Collection and extraction of occupational air samples for analysis of fungal DNA. (cdc.gov)
Epidemiology1
- Oilseed rape residues are a crucial determinant of stem canker epidemiology, as they support the sexual reproduction of the fungal pathogen Leptosphaeria maculans . (biorxiv.org)
Closely1
- Some recently discovered fungal viruses are most closely related to viruses that were thought to infect plants only. (theconversation.com)